

Single-cell deconvolution reveals high lineage- and location-dependent heterogeneity in mesenchymal multivisceral stage 4 colorectal cancer
- PMID: 38153787
- PMCID: PMC10904044
- DOI: 10.1172/JCI169576
Single-cell deconvolution reveals high lineage- and location-dependent heterogeneity in mesenchymal multivisceral stage 4 colorectal cancer
Abstract
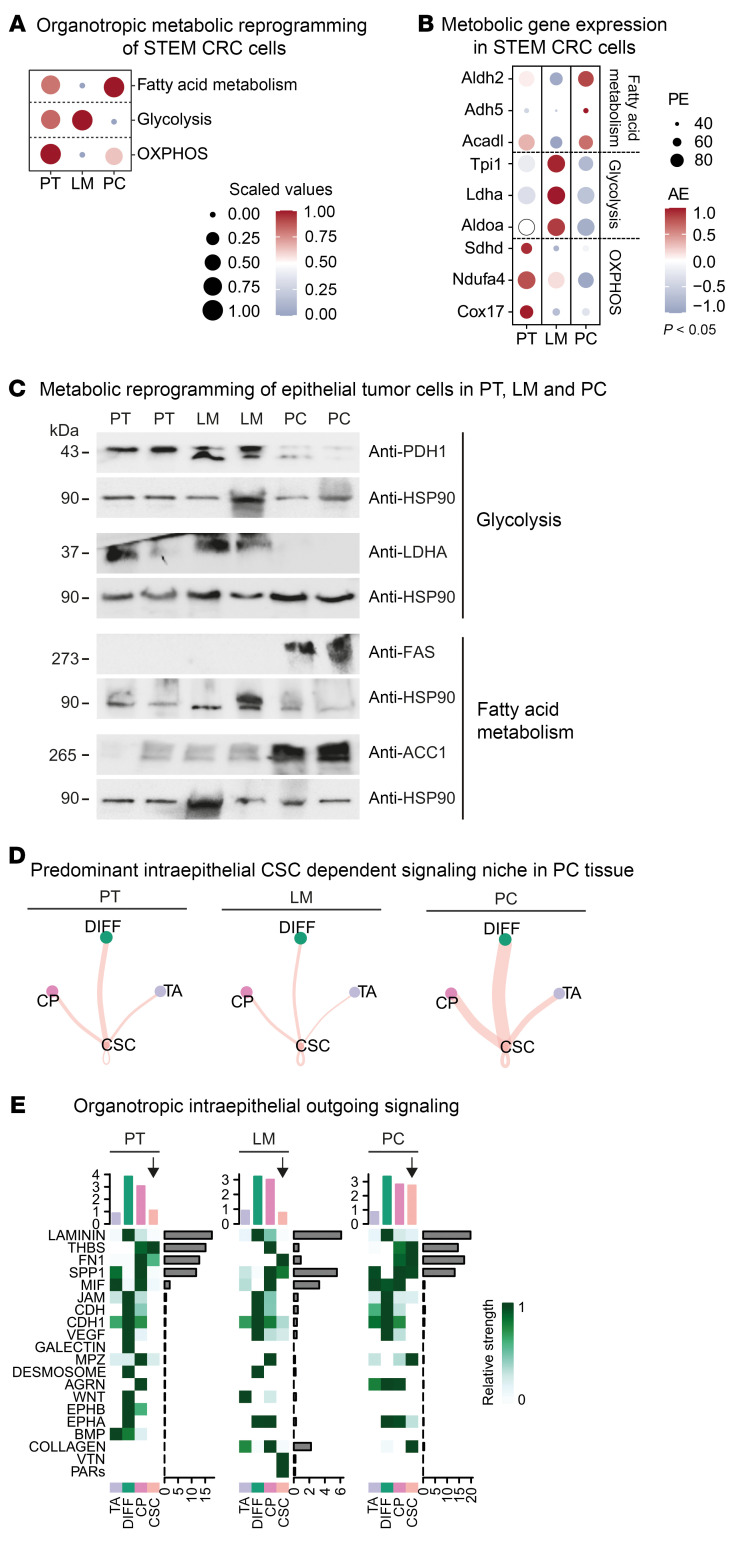
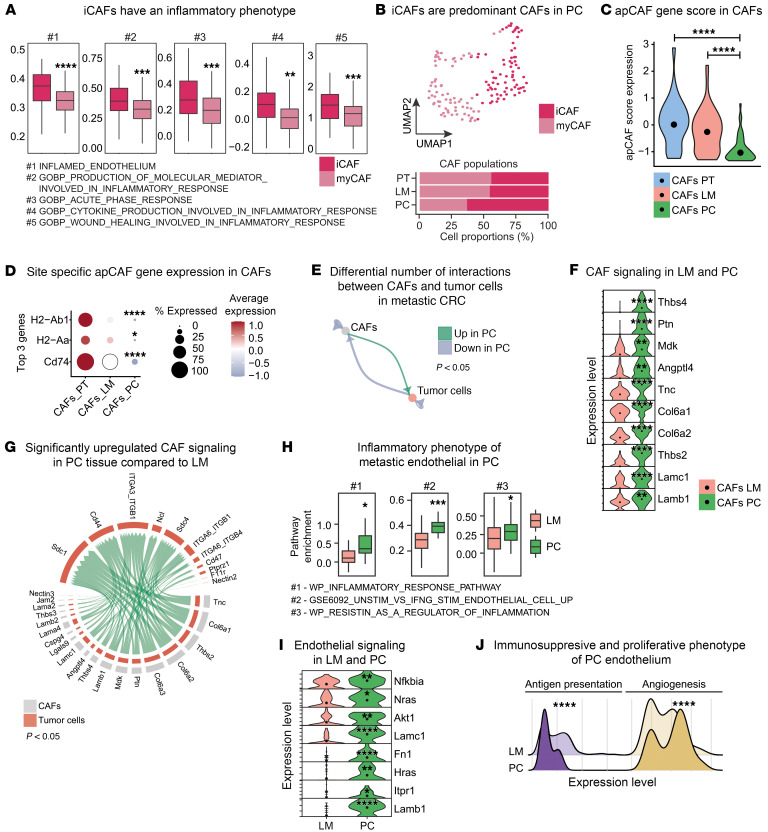
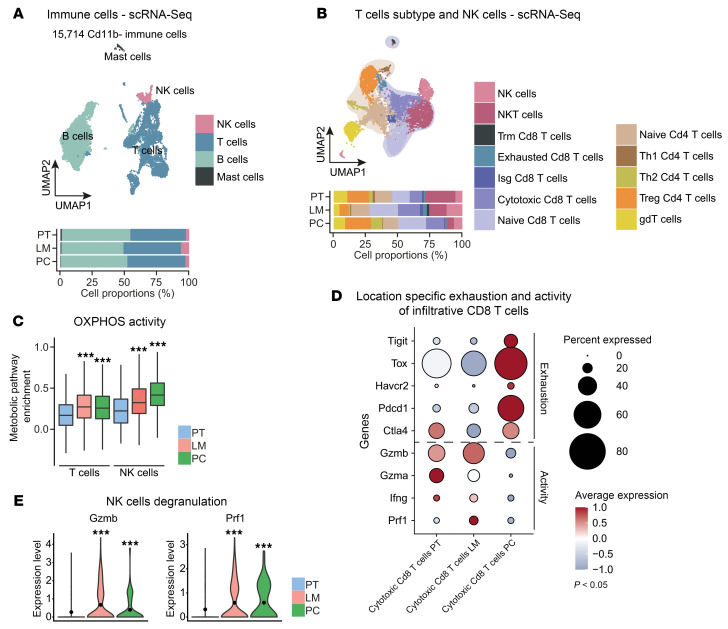
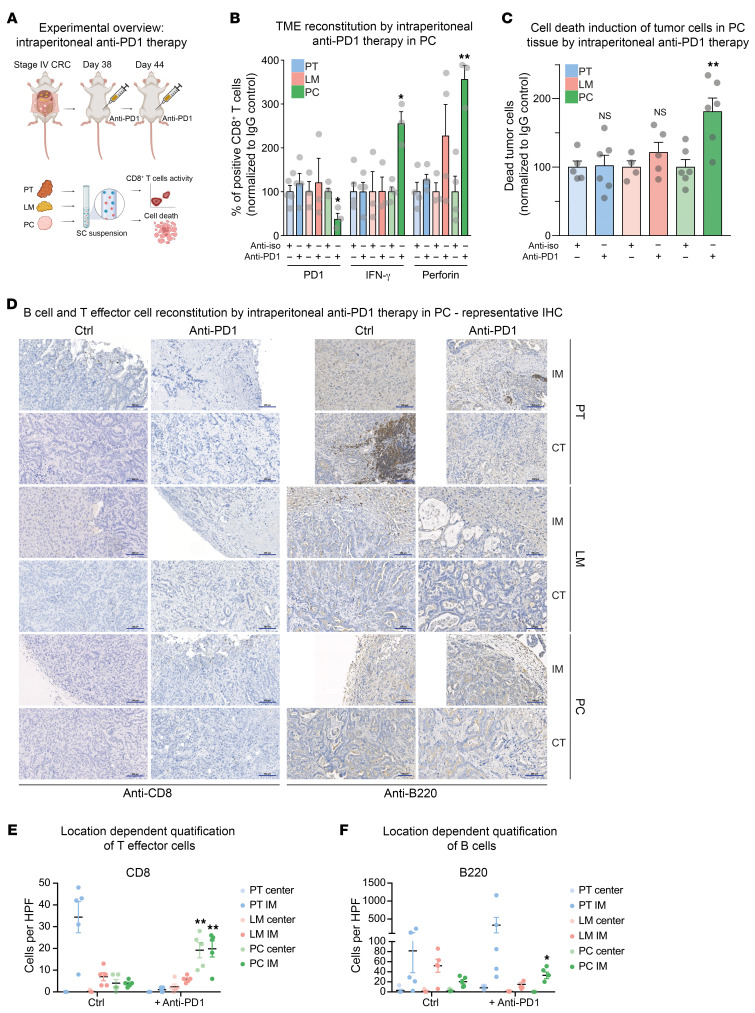
Metastasized colorectal cancer (CRC) is associated with a poor prognosis and rapid disease progression. Besides hepatic metastasis, peritoneal carcinomatosis is the major cause of death in Union for International Cancer Control (UICC) stage IV CRC patients. Insights into differential site-specific reconstitution of tumor cells and the corresponding tumor microenvironment are still missing. Here, we analyzed the transcriptome of single cells derived from murine multivisceral CRC and delineated the intermetastatic cellular heterogeneity regarding tumor epithelium, stroma, and immune cells. Interestingly, we found an intercellular site-specific network of cancer-associated fibroblasts and tumor epithelium during peritoneal metastasis as well as an autologous feed-forward loop in cancer stem cells. We furthermore deciphered a metastatic dysfunctional adaptive immunity by a loss of B cell-dependent antigen presentation and consecutive effector T cell exhaustion. Furthermore, we demonstrated major similarities of this murine metastatic CRC model with human disease and - based on the results of our analysis - provided an auspicious site-specific immunomodulatory treatment approach for stage IV CRC by intraperitoneal checkpoint inhibition.
Keywords: Cancer immunotherapy; Cell biology; Colorectal cancer; Mouse models; Oncology.
Figures





Comment in
- The multivisceral landscape of colorectal cancer metastasis: Implications for targeted therapies doi: 10.1172/JCI178331
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
