

C/EBPβ-induced lymphoid-to-myeloid transdifferentiation emulates granulocyte-monocyte progenitor biology
- PMID: 38157851
- PMCID: PMC10828814
- DOI: 10.1016/j.stemcr.2023.11.011
C/EBPβ-induced lymphoid-to-myeloid transdifferentiation emulates granulocyte-monocyte progenitor biology
Abstract
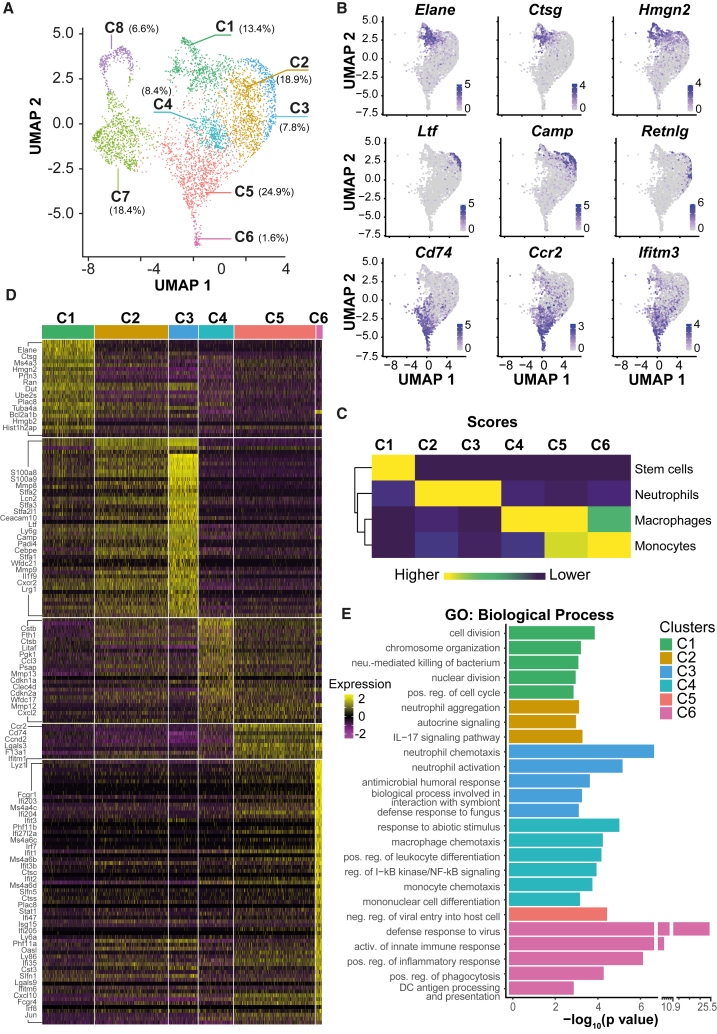
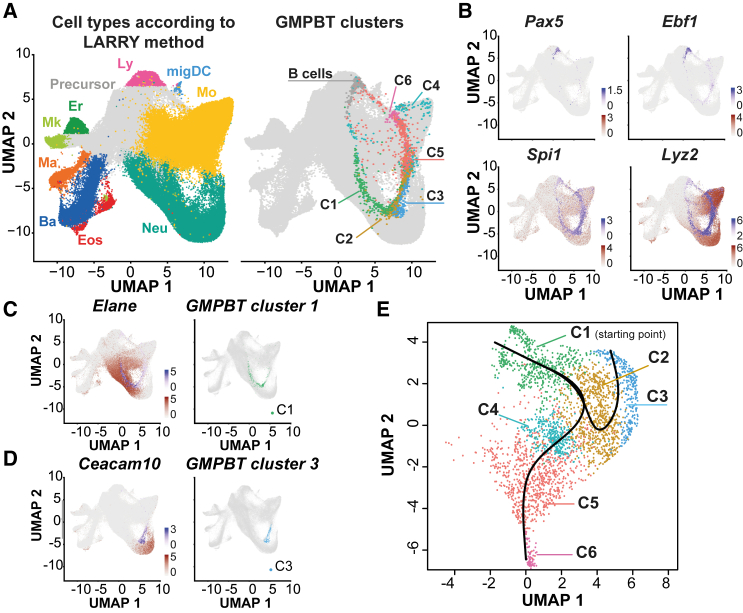
CCAAT/enhancer-binding protein beta (C/EBPβ) induces primary v-Abl immortalized mouse B cells to transdifferentiate (BT, B cell transdifferentiation) into granulocyte-macrophage progenitor-like cells (GMPBTs). GMPBTs maintain cytokine-independent self-renewal, lineage choice, and multilineage differentiation. Single-cell transcriptomics demonstrated that GMPBTs comprise a continuum of myelomonopoietic differentiation states that seamlessly fit into state-to-fate maps of normal granulocyte-macrophage progenitors (GMPs). Inactivating v-Abl kinase revealed the dependence on activated CSF2-JAK2-STAT5 signaling. Deleting IRF8 diminished monopoiesis and enhanced granulopoiesis while removing C/EBPβ-abrogated self-renewal and granulopoiesis but permitted macrophage differentiation. The GMPBT culture system is easily scalable to explore the basics of GMP biology and lineage commitment and largely reduces ethically and legislatively debatable, labor-intensive, and costly animal experiments.
Keywords: 3R principles; C/EBP; GMP; cell fate; granulocyte-macrophage progenitor; hematopoiesis; leukemia; myelopoiesis; transdifferentiation.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



Similar articles
-
C/EBP-Induced Transdifferentiation Reveals Granulocyte-Macrophage Precursor-like Plasticity of B Cells.Stem Cell Reports. 2017 Feb 14;8(2):346-359. doi: 10.1016/j.stemcr.2016.12.015. Epub 2017 Jan 19. Stem Cell Reports. 2017. PMID: 28111277 Free PMC article.
-
IRF8 acts in lineage-committed rather than oligopotent progenitors to control neutrophil vs monocyte production.Blood. 2015 Feb 26;125(9):1452-9. doi: 10.1182/blood-2014-09-600833. Epub 2015 Jan 16. Blood. 2015. PMID: 25597637
-
Granulopoiesis requires increased C/EBPα compared to monopoiesis, correlated with elevated Cebpa in immature G-CSF receptor versus M-CSF receptor expressing cells.PLoS One. 2014 Apr 21;9(4):e95784. doi: 10.1371/journal.pone.0095784. eCollection 2014. PLoS One. 2014. PMID: 24752325 Free PMC article.
-
Transcriptional control of granulocyte and monocyte development.Oncogene. 2007 Oct 15;26(47):6816-28. doi: 10.1038/sj.onc.1210764. Oncogene. 2007. PMID: 17934488 Review.
-
Regulation of granulocyte and monocyte differentiation by CCAAT/enhancer binding protein alpha.Blood Cells Mol Dis. 2003 Nov-Dec;31(3):338-41. doi: 10.1016/s1079-9796(03)00135-9. Blood Cells Mol Dis. 2003. PMID: 14636649 Review.
Cited by
-
Design and Evaluation of RNA Aptamer-Mediated Delivery of C/EBPβ siRNA for Oncological Therapy.J Nucleic Acids. 2025 Mar 3;2025:1461574. doi: 10.1155/jna/1461574. eCollection 2025. J Nucleic Acids. 2025. PMID: 40226120 Free PMC article.
-
Neutrophils in cancer: from biology to therapy.Cell Mol Immunol. 2025 Jan;22(1):4-23. doi: 10.1038/s41423-024-01244-9. Epub 2024 Dec 9. Cell Mol Immunol. 2025. PMID: 39653768 Free PMC article. Review.
-
Arginine methylation of the p30 C/EBPα oncoprotein regulates progenitor proliferation and myeloid differentiation.iScience. 2024 Oct 18;27(11):111199. doi: 10.1016/j.isci.2024.111199. eCollection 2024 Nov 15. iScience. 2024. PMID: 39555410 Free PMC article.
References
-
- Arendt D., Musser J.M., Baker C.V.H., Bergman A., Cepko C., Erwin D.H., Pavlicev M., Schlosser G., Widder S., Laubichler M.D., Wagner G.P. The origin and evolution of cell types. Nat. Rev. Genet. 2016;17:744–757. - PubMed
-
- Becher B., Tugues S., Greter M. GM-CSF: From Growth Factor to Central Mediator of Tissue Inflammation. Immunity. 2016;45:963–973. - PubMed
-
- Bussmann L.H., Schubert A., Vu Manh T.P., De Andres L., Desbordes S.C., Parra M., Zimmermann T., Rapino F., Rodriguez-Ubreva J., Ballestar E., Graf T. A robust and highly efficient immune cell reprogramming system. Cell Stem Cell. 2009;5:554–566. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
