

Astrocytes Drive Divergent Metabolic Gene Expression in Humans and Chimpanzees
- PMID: 38159045
- PMCID: PMC10829071
- DOI: 10.1093/gbe/evad239
Astrocytes Drive Divergent Metabolic Gene Expression in Humans and Chimpanzees
Abstract
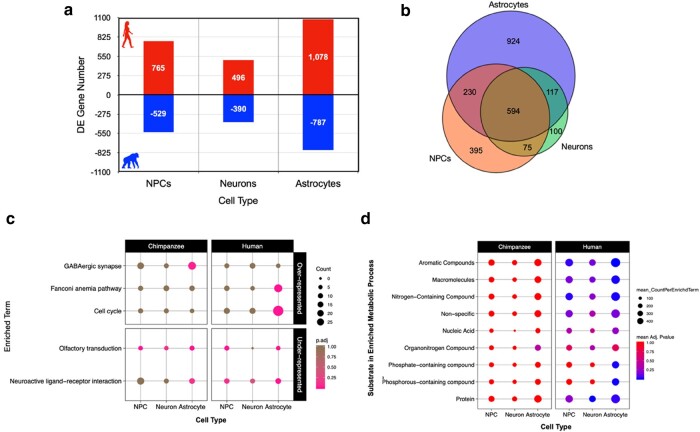
The human brain utilizes ∼20% of all of the body's metabolic resources, while chimpanzee brains use <10%. Although previous work shows significant differences in metabolic gene expression between the brains of primates, we have yet to fully resolve the contribution of distinct brain cell types. To investigate cell type-specific interspecies differences in brain gene expression, we conducted RNA-seq on neural progenitor cells, neurons, and astrocytes generated from induced pluripotent stem cells from humans and chimpanzees. Interspecies differential expression analyses revealed that twice as many genes exhibit differential expression in astrocytes (12.2% of all genes expressed) than neurons (5.8%). Pathway enrichment analyses determined that astrocytes, rather than neurons, diverged in expression of glucose and lactate transmembrane transport, as well as pyruvate processing and oxidative phosphorylation. These findings suggest that astrocytes may have contributed significantly to the evolution of greater brain glucose metabolism with proximity to humans.
Keywords: astrocytes; brain; evolution; genomics; metabolism; neurons.
© The Author(s) 2023. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures




References
-
- Aiello LC, Wheeler P. The expensive-tissue hypothesis: the brain and the digestive system in human and primate evolution. Curr Anthropol. 1995:36(2):199–221. 10.1086/204350. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
