

2-Bromopalmitate inhibits malignant behaviors of HPSCC cells by hindering the membrane location of Ras protein
- PMID: 38159074
- PMCID: PMC10903252
- DOI: 10.1177/15353702231220671
2-Bromopalmitate inhibits malignant behaviors of HPSCC cells by hindering the membrane location of Ras protein
Abstract
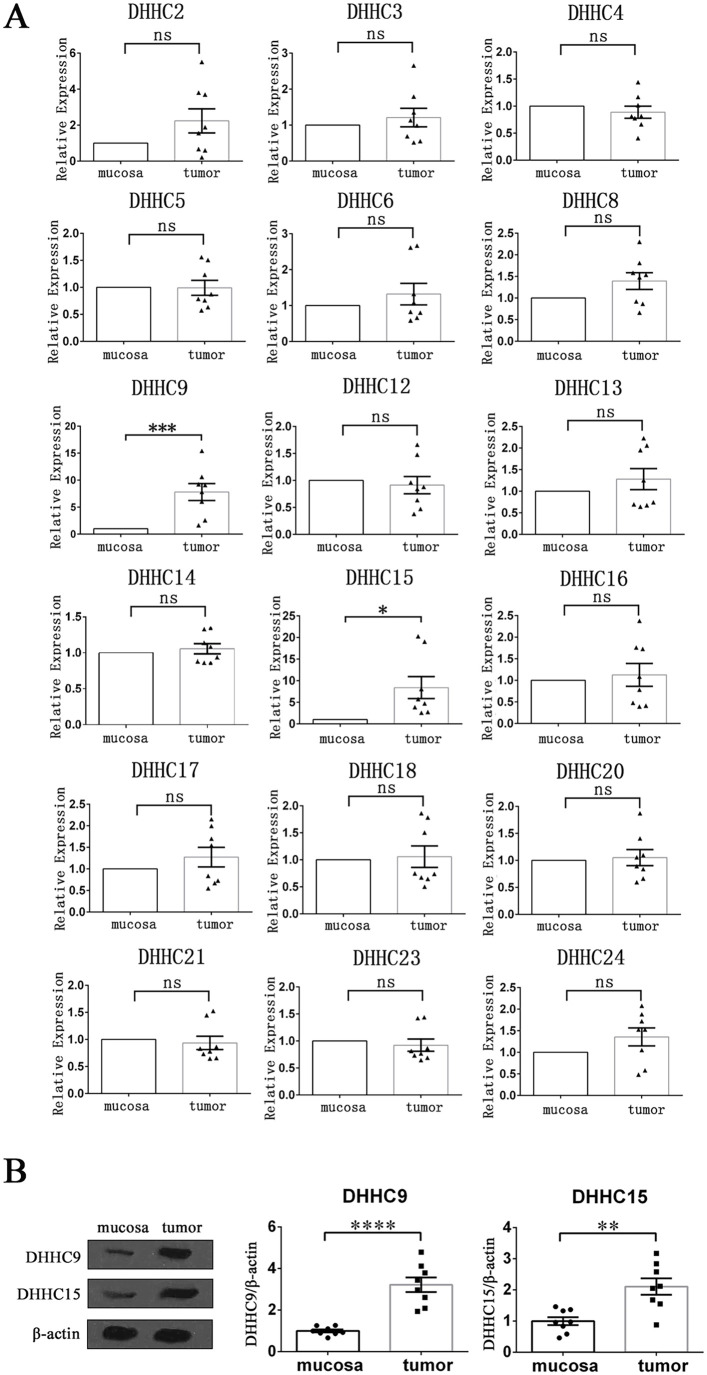
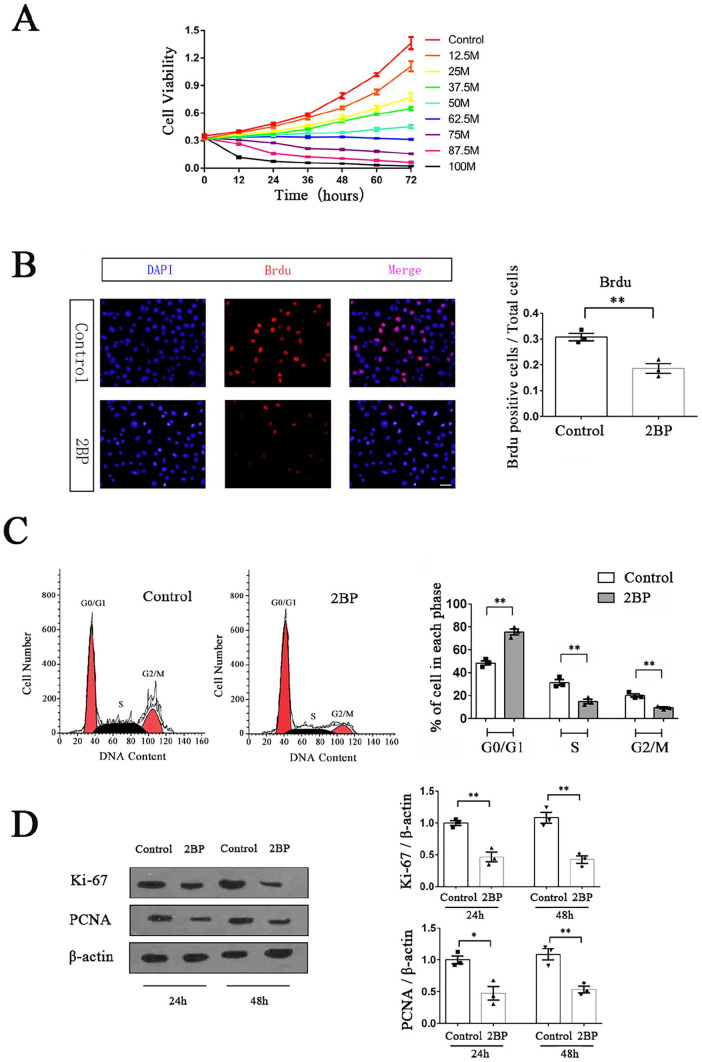
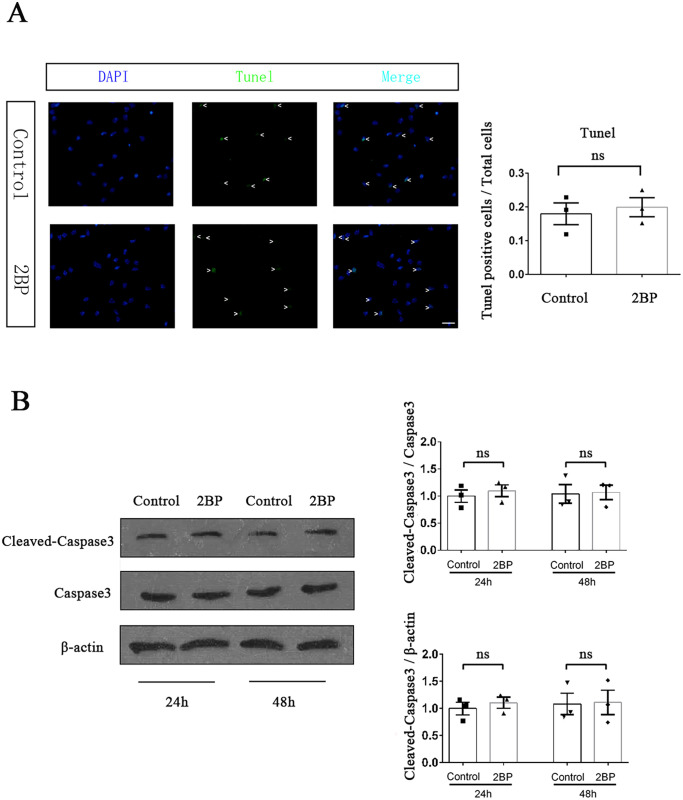
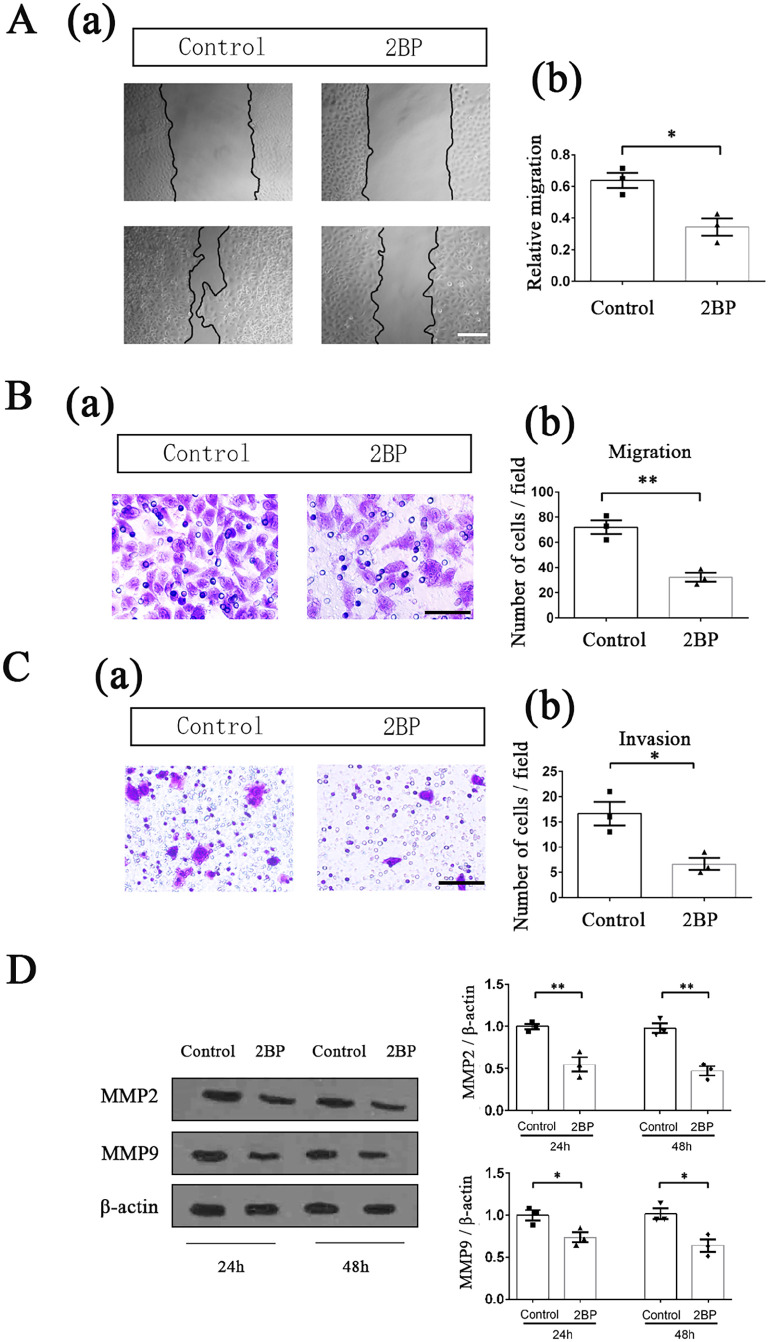
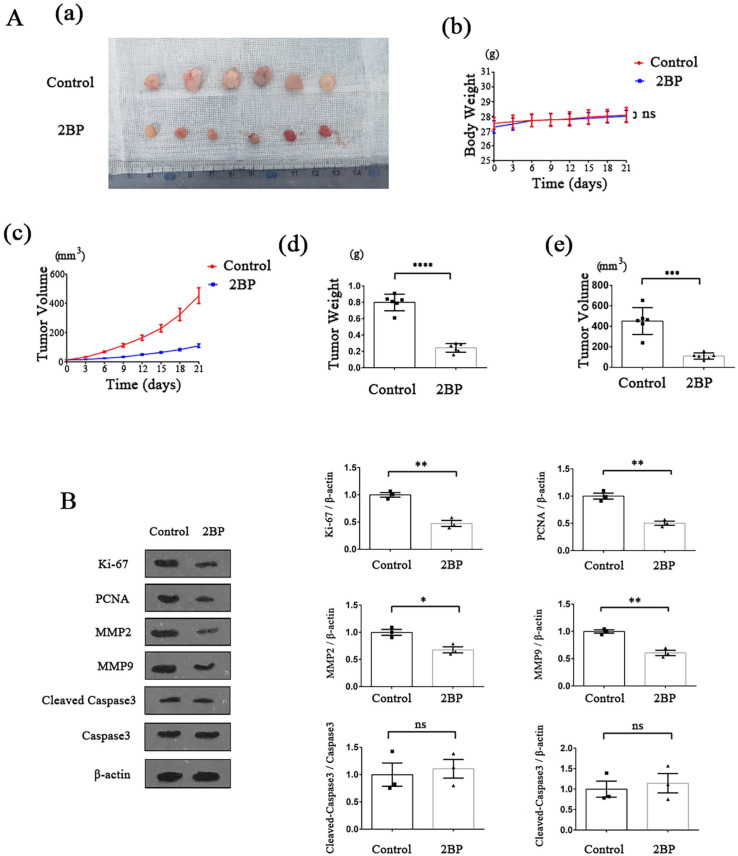
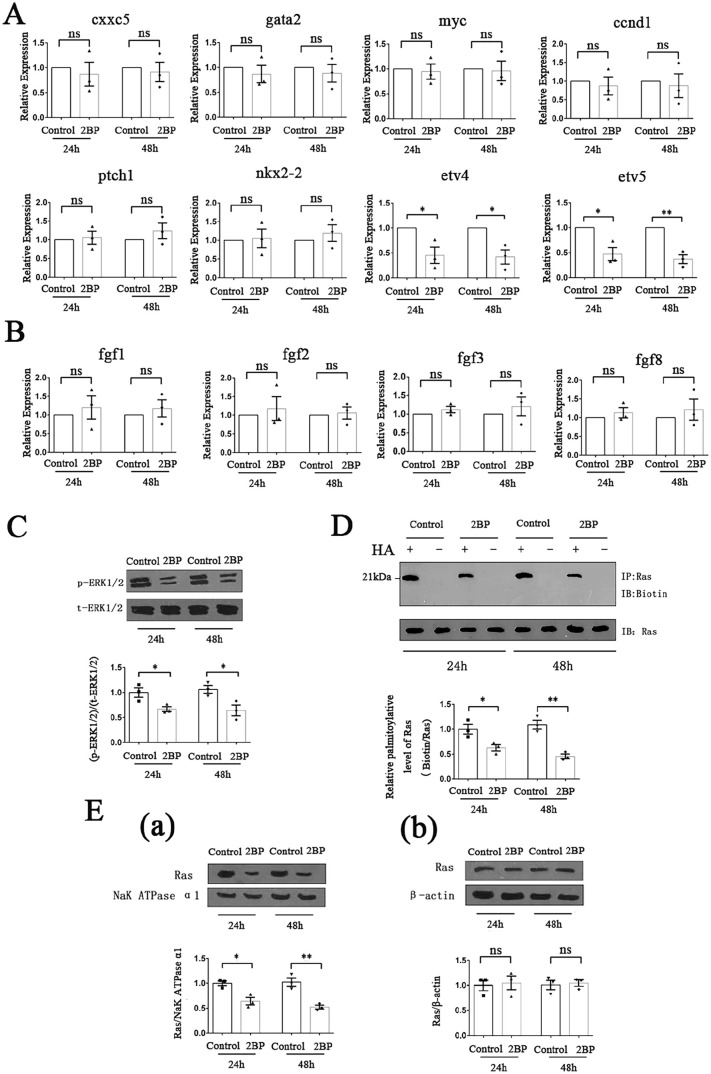
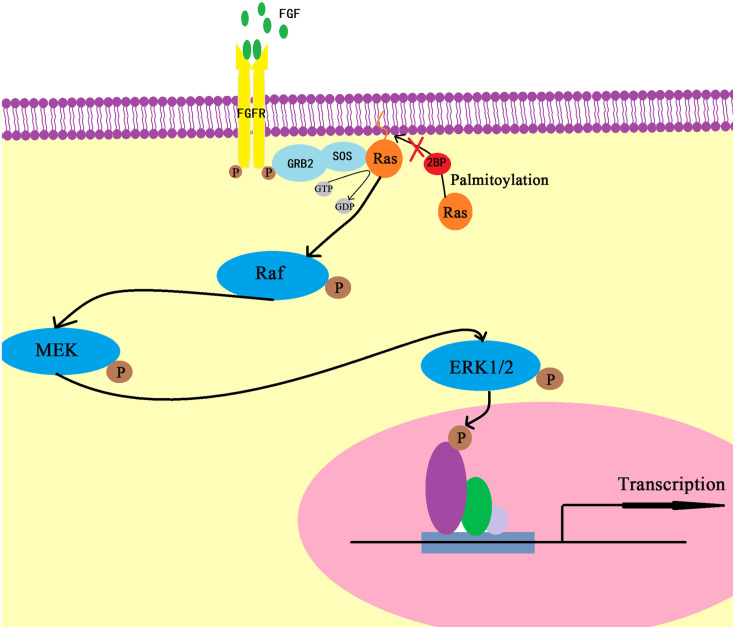
Palmitoylation, which is mediated by protein acyltransferase (PAT) and performs important biological functions, is the only reversible lipid modification in organism. To study the effect of protein palmitoylation on hypopharyngeal squamous cell carcinoma (HPSCC), the expression levels of 23 PATs in tumor tissues of 8 HPSCC patients were determined, and high mRNA and protein levels of DHHC9 and DHHC15 were found. Subsequently, we investigated the effect of 2-bromopalmitate (2BP), a small-molecular inhibitor of protein palmitoylation, on the behavior of Fadu cells in vitro (50 μM) and in nude mouse xenograft models (50 μmol/kg), and found that 2BP suppressed the proliferation, invasion, and migration of Fadu cells without increasing cell apoptosis. Mechanistically, the effect of 2BP on the transduction of BMP, Wnt, Shh, and FGF signaling pathways was tested with qRT-PCR, and its drug target was explored with western blotting and acyl-biotinyl exchange assay. Our results showed that 2BP inhibited the transduction of the FGF/ERK signaling pathway. The palmitoylation level of Ras protein decreased after 2BP treatment, and its distribution in the cell membrane structure was reduced significantly. The findings of this work reveal that protein palmitoylation mediated by DHHC9 and DHHC15 may play important roles in the occurrence and development of HPSCC. 2BP is able to inhibit the malignant biological behaviors of HPSCC cells, possibly via hindering the palmitoylation and membrane location of Ras protein, which might, in turn, offer a low-toxicity anti-cancer drug for targeting the treatment of HPSCC.
Keywords: 2-Bromopalmitate; FGF/ERK signaling pathway; Ras protein; hypopharyngeal squamous cell carcinoma; malignant behaviors; protein palmitoylation.
© 2023 by the Society for Experimental Biology and Medicine
Conflict of interest statement
Declaration of conflicting interestsThe author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous