

EZH2 inhibitors promote β-like cell regeneration in young and adult type 1 diabetes donors
- PMID: 38161208
- PMCID: PMC10757994
- DOI: 10.1038/s41392-023-01707-x
EZH2 inhibitors promote β-like cell regeneration in young and adult type 1 diabetes donors
Abstract
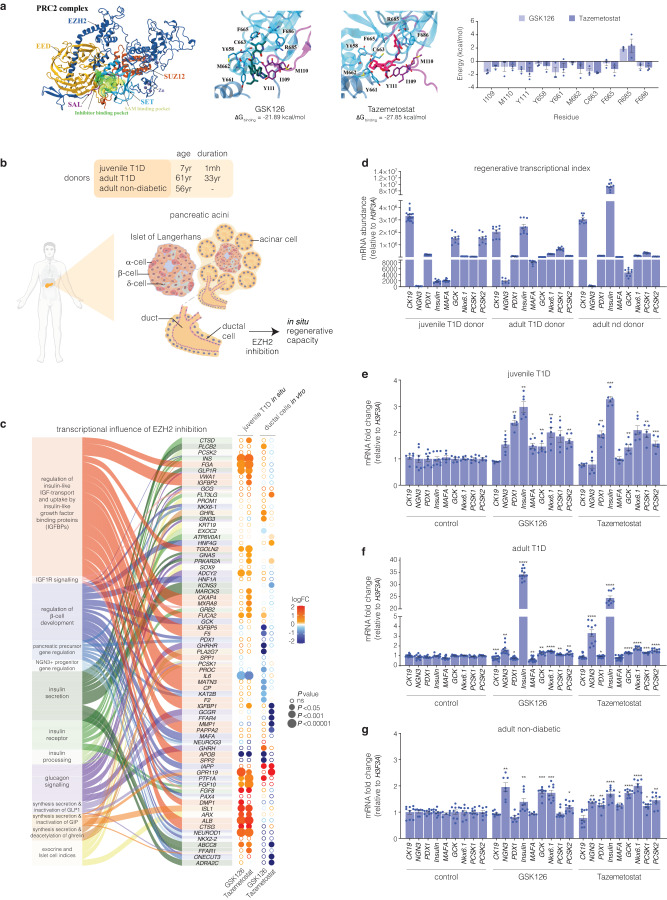
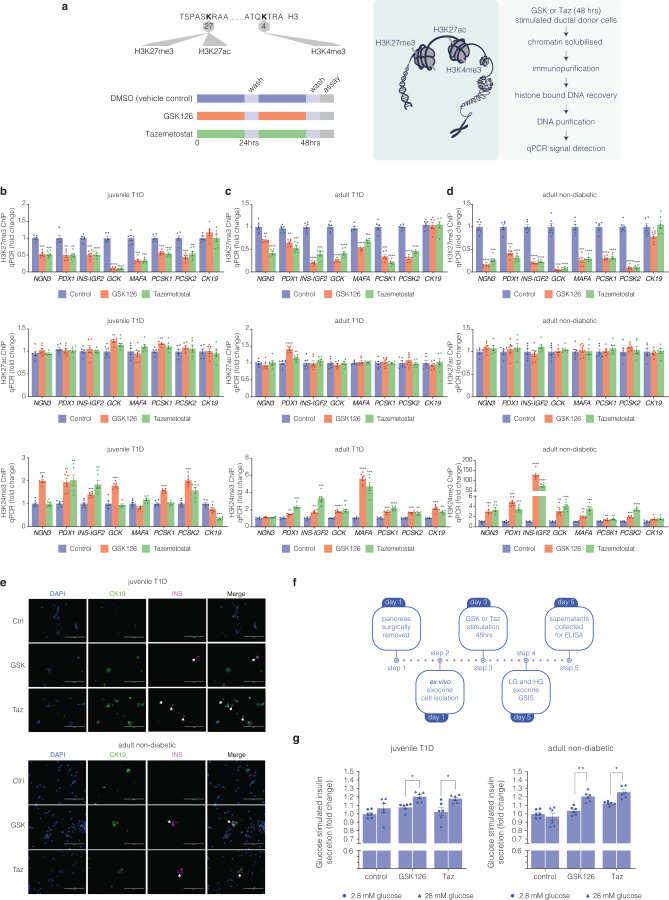
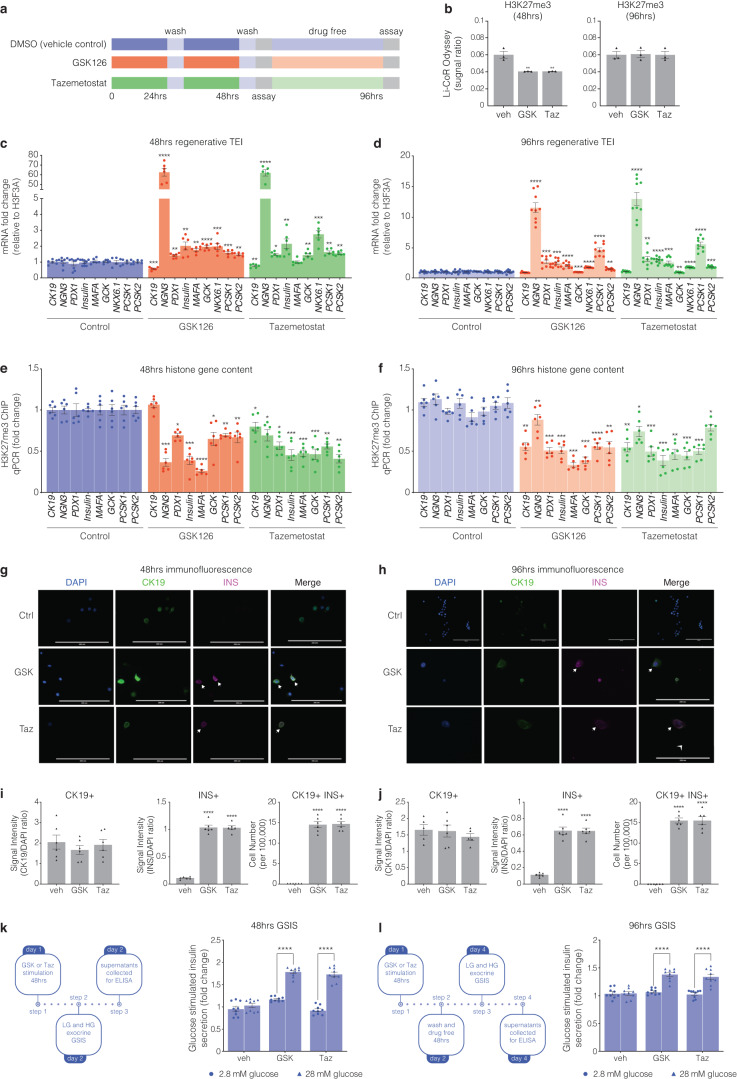
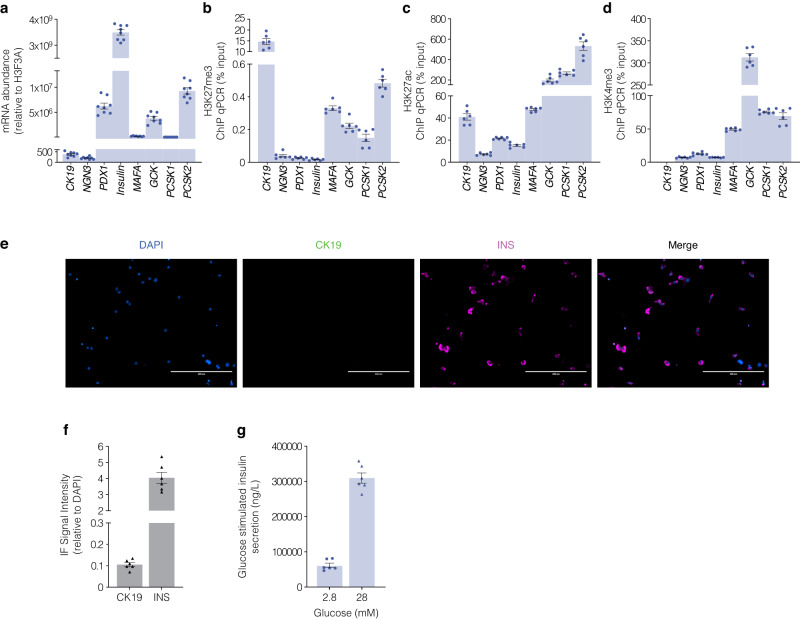
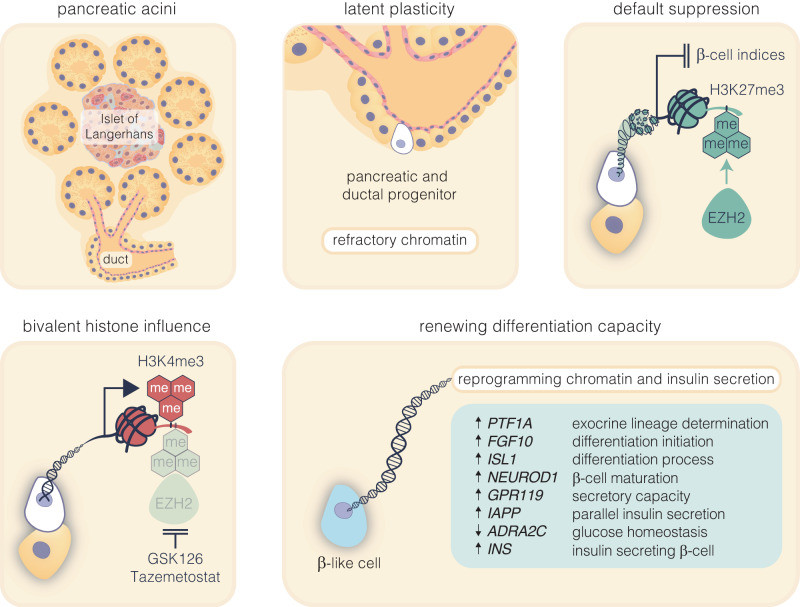
β-cells are a type of endocrine cell found in pancreatic islets that synthesize, store and release insulin. In type 1 diabetes (T1D), T-cells of the immune system selectively destroy the insulin-producing β-cells. Destruction of these cells leads to a lifelong dependence on exogenous insulin administration for survival. Consequently, there is an urgent need to identify novel therapies that stimulate β-cell growth and induce β-cell function. We and others have shown that pancreatic ductal progenitor cells are a promising source for regenerating β-cells for T1D owing to their inherent differentiation capacity. Default transcriptional suppression is refractory to exocrine reaction and tightly controls the regenerative potential by the EZH2 methyltransferase. In the present study, we show that transient stimulation of exocrine cells, derived from juvenile and adult T1D donors to the FDA-approved EZH2 inhibitors GSK126 and Tazemetostat (Taz) influence a phenotypic shift towards a β-like cell identity. The transition from repressed to permissive chromatin states are dependent on bivalent H3K27me3 and H3K4me3 chromatin modification. Targeting EZH2 is fundamental to β-cell regenerative potential. Reprogrammed pancreatic ductal cells exhibit insulin production and secretion in response to a physiological glucose challenge ex vivo. These pre-clinical studies underscore the potential of small molecule inhibitors as novel modulators of ductal progenitor differentiation and a promising new approach for the restoration of β-like cell function.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Inhibition of pancreatic EZH2 restores progenitor insulin in T1D donor.Signal Transduct Target Ther. 2022 Jul 22;7(1):248. doi: 10.1038/s41392-022-01034-7. Signal Transduct Target Ther. 2022. PMID: 35864094 Free PMC article.
-
Pharmacological inhibition of human EZH2 can influence a regenerative β-like cell capacity with in vitro insulin release in pancreatic ductal cells.Clin Epigenetics. 2023 Jun 12;15(1):101. doi: 10.1186/s13148-023-01491-z. Clin Epigenetics. 2023. PMID: 37309004 Free PMC article.
-
Modulation of the pancreatic islet-stress axis as a novel potential therapeutic target in diabetes mellitus.Vitam Horm. 2014;95:195-222. doi: 10.1016/B978-0-12-800174-5.00008-9. Vitam Horm. 2014. PMID: 24559919 Review.
-
Dynamics of genomic H3K27me3 domains and role of EZH2 during pancreatic endocrine specification.EMBO J. 2014 Oct 1;33(19):2157-70. doi: 10.15252/embj.201488671. Epub 2014 Aug 8. EMBO J. 2014. PMID: 25107471 Free PMC article.
-
Can pancreatic duct-derived progenitors be a source of islet regeneration?Biochem Biophys Res Commun. 2009 Jun 12;383(4):383-5. doi: 10.1016/j.bbrc.2009.03.114. Epub 2009 Mar 24. Biochem Biophys Res Commun. 2009. PMID: 19324022 Review.
Cited by
-
Advancing type 1 diabetes therapy: autologous islet transplant breakthrough.Signal Transduct Target Ther. 2024 Dec 23;9(1):366. doi: 10.1038/s41392-024-02090-x. Signal Transduct Target Ther. 2024. PMID: 39710778 Free PMC article. No abstract available.
-
The role of endothelial cells in pancreatic islet development, transplantation and culture.Front Cell Dev Biol. 2025 Apr 22;13:1558137. doi: 10.3389/fcell.2025.1558137. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40330424 Free PMC article. Review.
-
Epigenetics Plays a Role in the Pathogenesis and Treatment of Diabetes.Genes (Basel). 2025 Jun 30;16(7):769. doi: 10.3390/genes16070769. Genes (Basel). 2025. PMID: 40725425 Free PMC article. Review.
-
Islet Cell Replacement and Regeneration for Type 1 Diabetes: Current Developments and Future Prospects.BioDrugs. 2025 Mar;39(2):261-280. doi: 10.1007/s40259-025-00703-7. Epub 2025 Feb 7. BioDrugs. 2025. PMID: 39918671 Free PMC article. Review.
-
Targeting EZH2 in autoimmune diseases: unraveling epigenetic regulation and therapeutic potential.Naunyn Schmiedebergs Arch Pharmacol. 2025 Apr 8. doi: 10.1007/s00210-025-04127-6. Online ahead of print. Naunyn Schmiedebergs Arch Pharmacol. 2025. PMID: 40198399 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
