

Reappraisal of the DNA phosphorothioate modification machinery: uncovering neglected functional modalities and identification of new counter-invader defense systems
- PMID: 38163645
- PMCID: PMC10853773
- DOI: 10.1093/nar/gkad1213
Reappraisal of the DNA phosphorothioate modification machinery: uncovering neglected functional modalities and identification of new counter-invader defense systems
Abstract
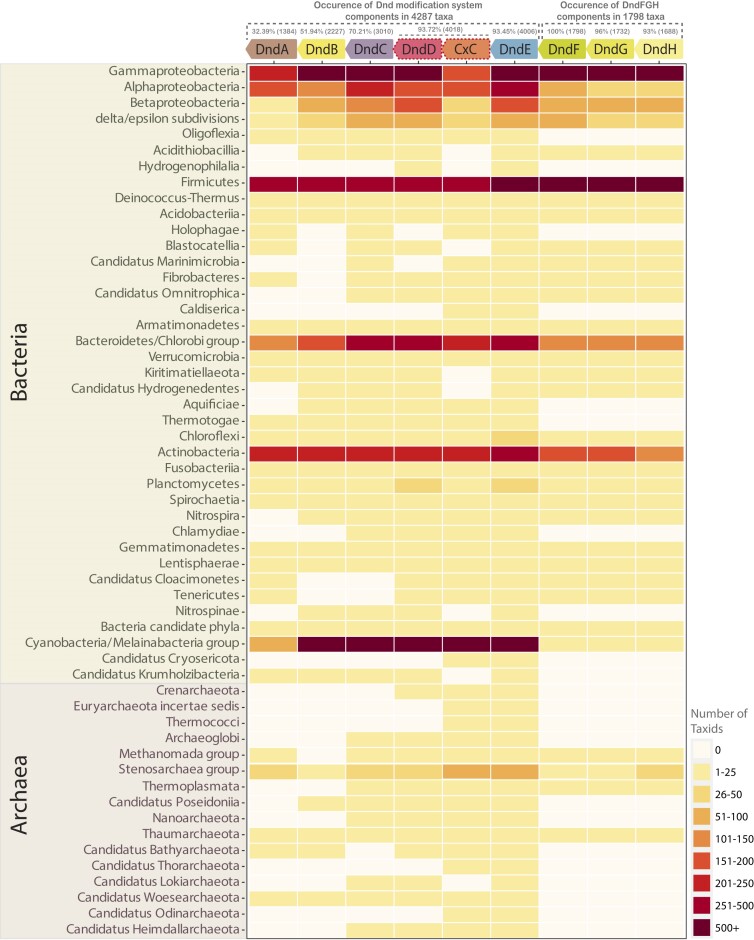
The DndABCDE systems catalysing the unusual phosphorothioate (PT) DNA backbone modification, and the DndFGH systems, which restrict invasive DNA, have enigmatic and paradoxical features. Using comparative genomics and sequence-structure analyses, we show that the DndABCDE module is commonly functionally decoupled from the DndFGH module. However, the modification gene-neighborhoods encode other nucleases, potentially acting as the actual restriction components or suicide effectors limiting propagation of the selfish elements. The modification module's core consists of a coevolving gene-pair encoding the DNA-scanning apparatus - a DndD/CxC-clade ABC ATPase and DndE with two ribbon-helix-helix (MetJ/Arc) DNA-binding domains. Diversification of DndE's DNA-binding interface suggests a multiplicity of target specificities. Additionally, many systems feature DNA cytosine methylase genes instead of PT modification, indicating the DndDE core can recruit other nucleobase modifications. We show that DndFGH is a distinct counter-invader system with several previously uncharacterized domains, including a nucleotide kinase. These likely trigger its restriction endonuclease domain in response to multiple stimuli, like nucleotides, while blocking protective modifications by invader methylases. Remarkably, different DndH variants contain a HerA/FtsK ATPase domain acquired from multiple sources, including cellular genome-segregation systems and mobile elements. Thus, we uncovered novel HerA/FtsK-dependent defense systems that might intercept invasive DNA during replication, conjugation, or packaging.
Plain language summary
Bacteria defend against selfish genetic elements by distinguishing their genetic material through special chemical modifications and using specific enzymes to break down viral DNA. This study explores the Dnd defense system, revealing several of its poorly understood facets. The Dnd modification system, utilizing sulfur to distinguish bacterial from viral DNA, cooperates with various anti-viral and cell-suicide nuclease enzymes to limit viral infection. While previously considered its restriction component, DndFGH emerges as an independent defense system, recognizing signals like nucleotides and DNA to thwart protective modifications of invader DNA. DndH, featuring diverse versions of the HerA/FtsK ATPase domain, helped unveil several unrecognized bacterial defense systems. This discovery illuminates sophisticated bacterial defenses against viral threats during crucial cellular processes.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures








Similar articles
-
Comparative genomics of the FtsK-HerA superfamily of pumping ATPases: implications for the origins of chromosome segregation, cell division and viral capsid packaging.Nucleic Acids Res. 2004 Oct 5;32(17):5260-79. doi: 10.1093/nar/gkh828. Print 2004. Nucleic Acids Res. 2004. PMID: 15466593 Free PMC article.
-
A DNA phosphorothioation-based Dnd defense system provides resistance against various phages and is compatible with the Ssp defense system.mBio. 2023 Aug 31;14(4):e0093323. doi: 10.1128/mbio.00933-23. Epub 2023 Jun 1. mBio. 2023. PMID: 37260233 Free PMC article.
-
Structural and Functional Analysis of DndE Involved in DNA Phosphorothioation in the Haloalkaliphilic Archaea Natronorubrum bangense JCM10635.mBio. 2022 Jun 28;13(3):e0071622. doi: 10.1128/mbio.00716-22. Epub 2022 Apr 14. mBio. 2022. PMID: 35420474 Free PMC article.
-
Comparative genomics and evolutionary trajectories of viral ATP dependent DNA-packaging systems.Genome Dyn. 2007;3:48-65. doi: 10.1159/000107603. Genome Dyn. 2007. PMID: 18753784 Review.
-
DNA phosphorothioate modification-a new multi-functional epigenetic system in bacteria.FEMS Microbiol Rev. 2019 Mar 1;43(2):109-122. doi: 10.1093/femsre/fuy036. FEMS Microbiol Rev. 2019. PMID: 30289455 Free PMC article. Review.
Cited by
-
A DNA phosphorothioation pathway via adenylated intermediate modulates Tdp machinery.Nat Chem Biol. 2025 Aug;21(8):1160-1170. doi: 10.1038/s41589-024-01832-w. Epub 2025 Jan 16. Nat Chem Biol. 2025. PMID: 39820821
-
Temporal dynamics and metagenomics of phosphorothioate epigenomes in the human gut microbiome.Microbiome. 2025 Mar 24;13(1):81. doi: 10.1186/s40168-025-02071-4. Microbiome. 2025. PMID: 40128848 Free PMC article.
-
Aeromonas trota Is Highly Refractory to Acquire Exogenous Genetic Material.Microorganisms. 2024 May 28;12(6):1091. doi: 10.3390/microorganisms12061091. Microorganisms. 2024. PMID: 38930473 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
