

Bilateral Retinofugal Pathfinding Impairments Limit Behavioral Compensation in Near-Congenital One-Eyed Xenopus laevis
- PMID: 38164595
- PMCID: PMC10849038
- DOI: 10.1523/ENEURO.0371-23.2023
Bilateral Retinofugal Pathfinding Impairments Limit Behavioral Compensation in Near-Congenital One-Eyed Xenopus laevis
Abstract
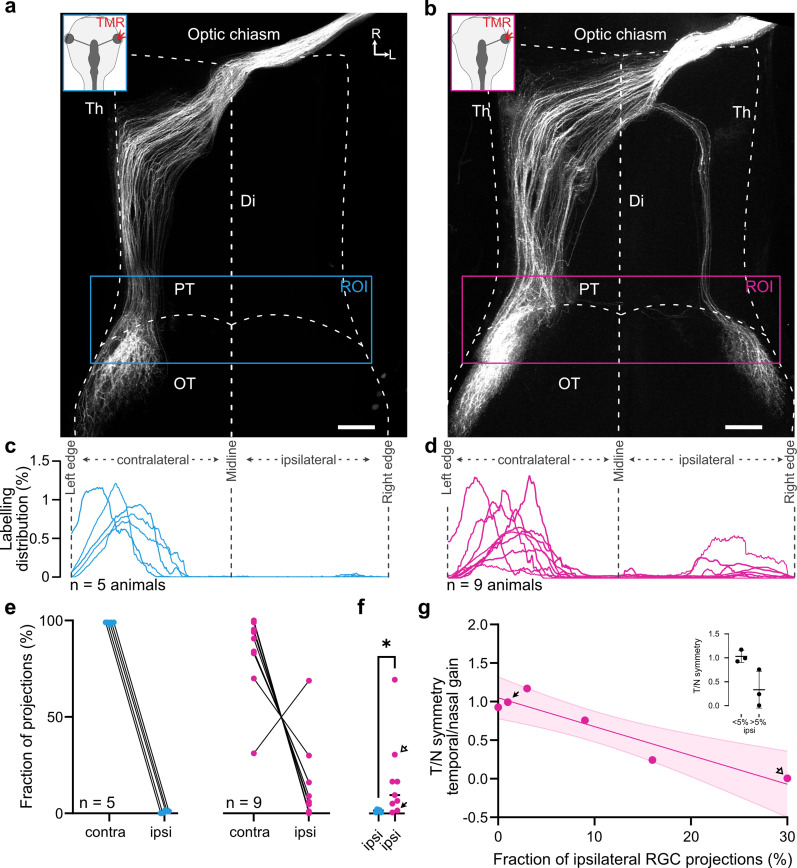
To generate a coherent visual percept, information from both eyes must be appropriately transmitted into the brain, where binocular integration forms the substrate for visuomotor behaviors. To establish the anatomical substrate for binocular integration, the presence of bilateral eyes and interaction of both optic nerves during retinotectal development play a key role. However, the extent to which embryonic monocularly derived visual circuits can convey visuomotor behaviors is unknown. In this study, we assessed the retinotectal anatomy and visuomotor performance of embryonically generated one-eyed tadpoles. In one-eyed animals, the axons of retinal ganglion cells from the singular remaining eye exhibited striking irregularities in their central projections in the brain, generating a noncanonical ipsilateral retinotectal projection. This data is indicative of impaired pathfinding abilities. We further show that these novel projections are correlated with an impairment of behavioral compensation for the loss of one eye.
Keywords: OKR; eye extirpation; frog; optokinetic reflex; plasticity; retinotectal.
Copyright © 2024 Forsthofer, Gordy et al.
Figures




Similar articles
-
Direct intertectal inputs are an integral component of the bilateral sensorimotor circuit for behavior in Xenopus tadpoles.J Neurophysiol. 2018 May 1;119(5):1947-1961. doi: 10.1152/jn.00051.2018. Epub 2018 Feb 14. J Neurophysiol. 2018. PMID: 29442555 Free PMC article.
-
Retinotectal map formation in dually innervated tecta: a regeneration study in Xenopus with one compound eye following bilateral optic nerve section.J Comp Neurol. 1982 Apr 1;206(2):119-30. doi: 10.1002/cne.902060203. J Comp Neurol. 1982. PMID: 7085924
-
Retinal specificity in eye fragments: investigations on the retinotectal projections of different quarter-eyes in Xenopus laevis.Exp Brain Res. 1994;102(2):272-86. doi: 10.1007/BF00227514. Exp Brain Res. 1994. PMID: 7705505
-
Plasticity in the tectum of Xenopus laevis: binocular maps.Prog Neurobiol. 1999 Oct;59(2):81-106. doi: 10.1016/s0301-0082(98)00096-3. Prog Neurobiol. 1999. PMID: 10463791 Review.
-
Early development and function of the Xenopus tadpole retinotectal circuit.Curr Opin Neurobiol. 2016 Dec;41:17-23. doi: 10.1016/j.conb.2016.07.002. Epub 2016 Jul 29. Curr Opin Neurobiol. 2016. PMID: 27475307 Review.
References
MeSH terms
LinkOut - more resources
Full Text Sources