

Dissolution of ribonucleoprotein condensates by the embryonic stem cell protein L1TD1
- PMID: 38165001
- PMCID: PMC11014241
- DOI: 10.1093/nar/gkad1244
Dissolution of ribonucleoprotein condensates by the embryonic stem cell protein L1TD1
Abstract
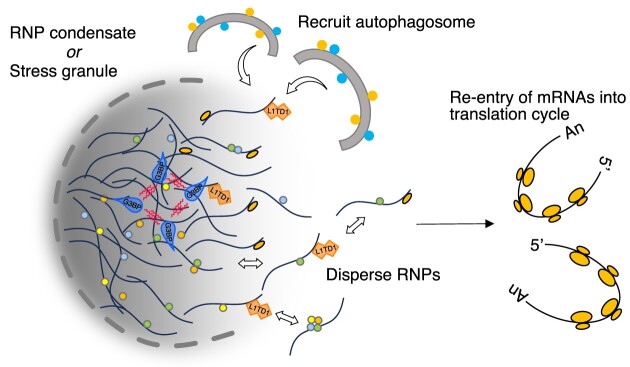
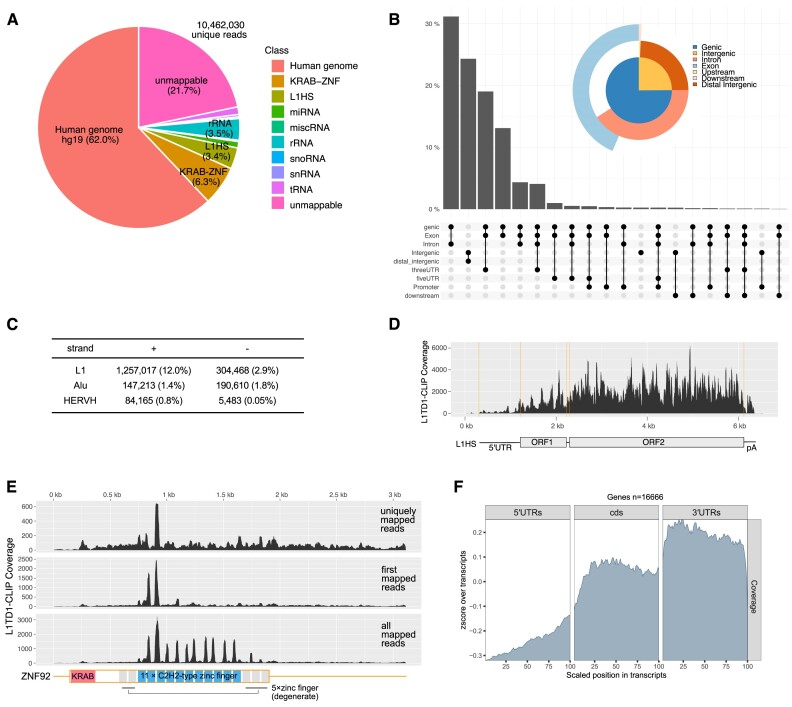
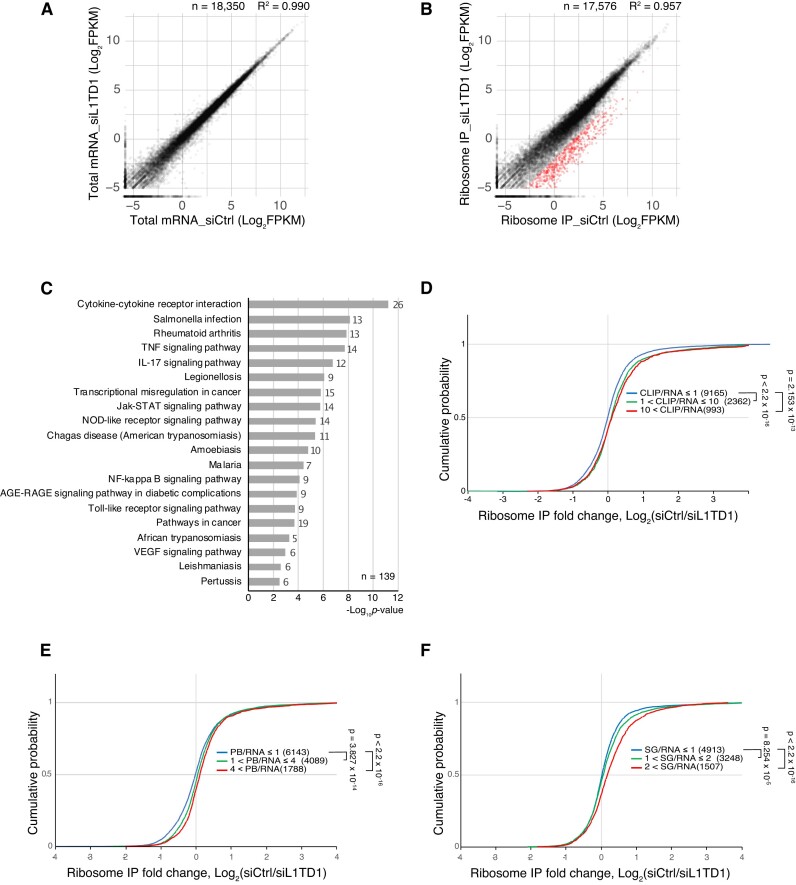
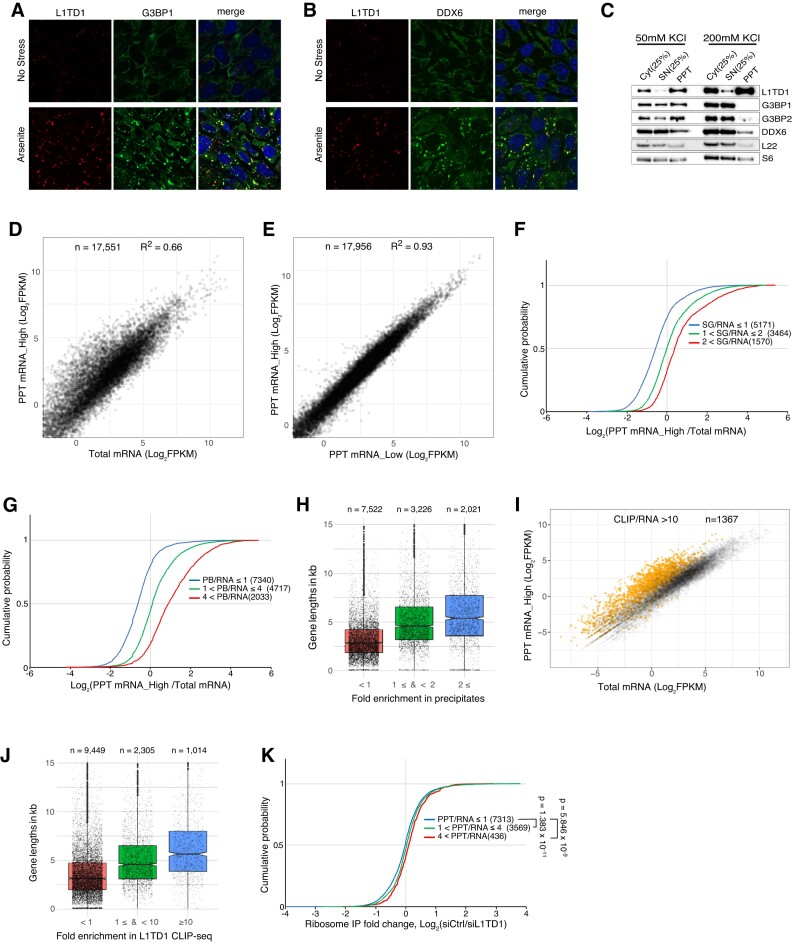
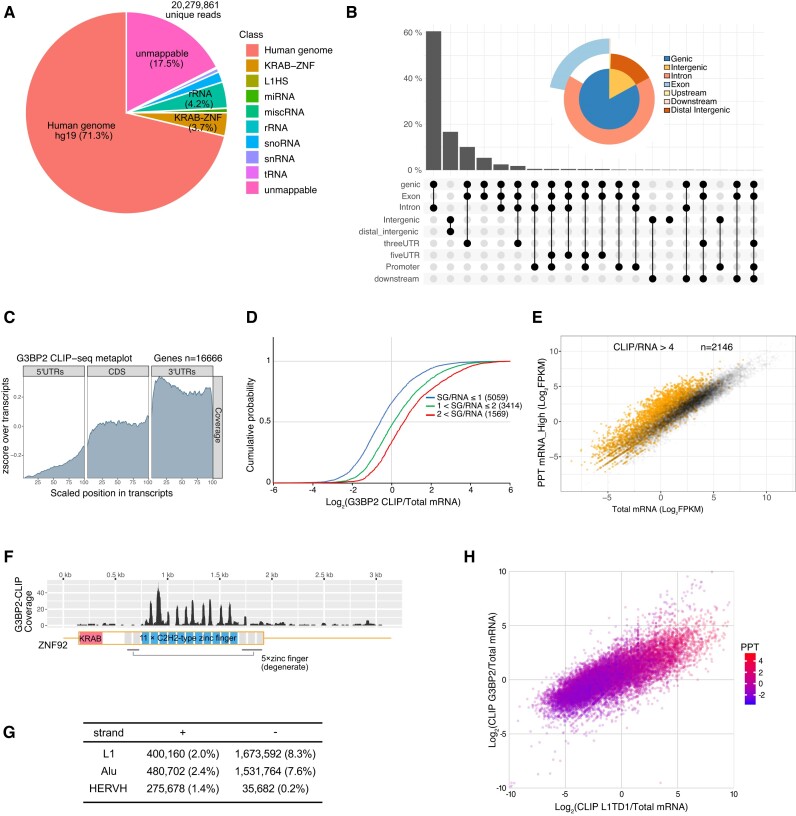
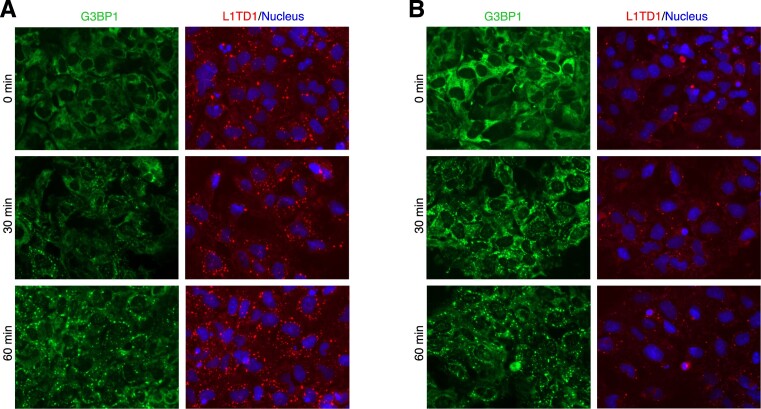
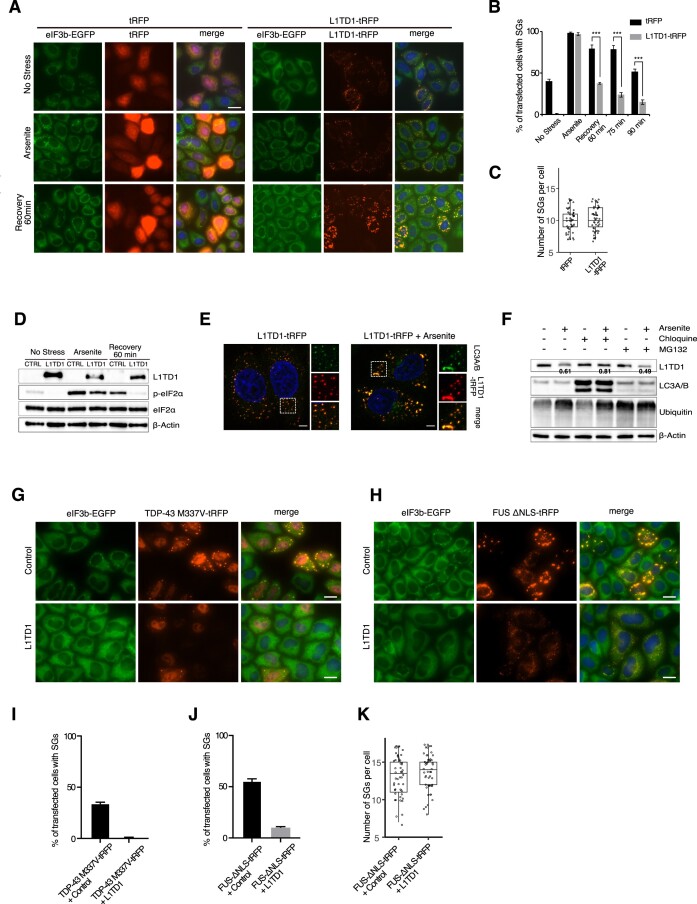
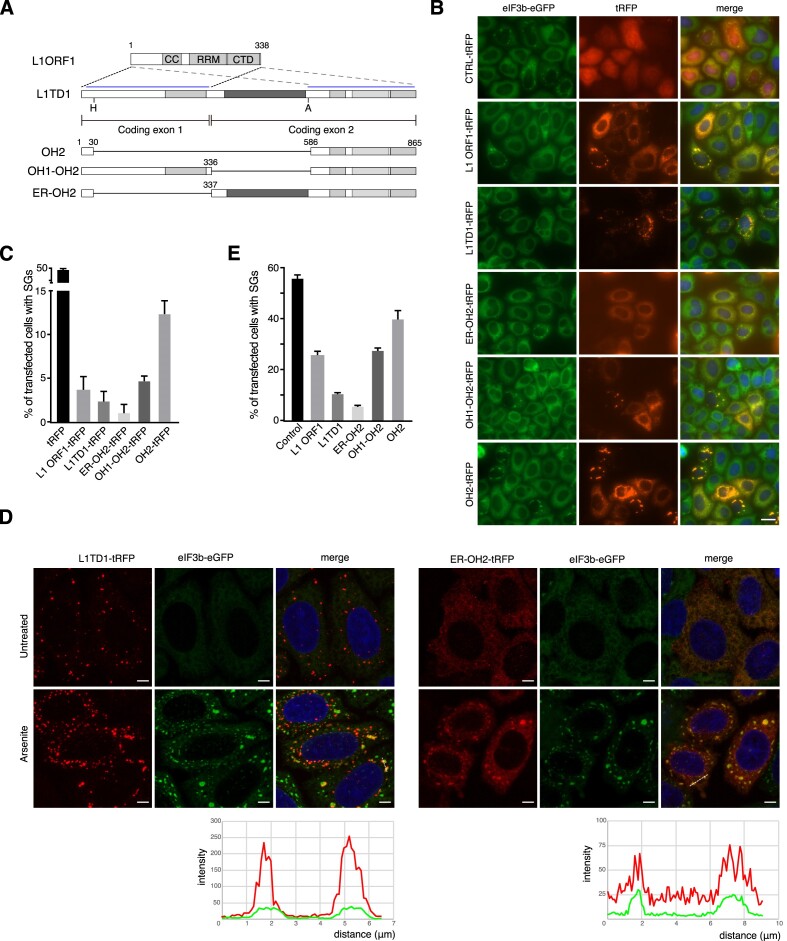
L1TD1 is a cytoplasmic RNA-binding protein specifically expressed in pluripotent stem cells and, unlike its mouse ortholog, is essential for the maintenance of stemness in human cells. Although L1TD1 is the only known protein-coding gene domesticated from a LINE-1 (L1) retroelement, the functional legacy of its ancestral protein, ORF1p of L1, and how it is manifested in L1TD1 are still unknown. Here, we determined RNAs associated with L1TD1 and found that, like ORF1p, L1TD1 binds L1 RNAs and localizes to high-density ribonucleoprotein (RNP) condensates. Unexpectedly, L1TD1 enhanced the translation of a subset of mRNAs enriched in the condensates. L1TD1 depletion promoted the formation of stress granules in embryonic stem cells. In HeLa cells, ectopically expressed L1TD1 facilitated the dissolution of stress granules and granules formed by pathological mutations of TDP-43 and FUS. The glutamate-rich domain and the ORF1-homology domain of L1TD1 facilitated dispersal of the RNPs and induced autophagy, respectively. These results provide insights into how L1TD1 regulates gene expression in pluripotent stem cells. We propose that the ability of L1TD1 to dissolve stress granules may provide novel opportunities for treatment of neurodegenerative diseases caused by disturbed stress granule dynamics.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Mitsui K., Tokuzawa Y., Itoh H., Segawa K., Murakami M., Takahashi K., Maruyama M., Maeda M., Yamanaka S.. The homeoprotein nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell. 2003; 113:631–642. - PubMed
-
- Närvä E., Rahkonen N., Emani M.R., Lund R., Pursiheimo J.-P., Nästi J., Autio R., Rasool O., Denessiouk K., Lähdesmäki H.et al. .. RNA-binding protein L1TD1 interacts with LIN28 via RNA and is required for Human embryonic stem cell self-renewal and cancer cell proliferation. Stem Cells. 2012; 30:452–460. - PMC - PubMed
-
- Santos M.C.T., Silva P.B.G., Rodini C.O., Furukawa G., Marco Antonio D.S., Zanotto-Filho A., Moreira J.C.F., Okamoto O.K.. Embryonic stem cell-related protein L1TD1 is required for cell viability, neurosphere formation, and chemoresistance in medulloblastoma. Stem Cells Dev. 2015; 24:2700–2708. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
