

THUMPD2 catalyzes the N2-methylation of U6 snRNA of the spliceosome catalytic center and regulates pre-mRNA splicing and retinal degeneration
- PMID: 38165050
- PMCID: PMC11014329
- DOI: 10.1093/nar/gkad1243
THUMPD2 catalyzes the N2-methylation of U6 snRNA of the spliceosome catalytic center and regulates pre-mRNA splicing and retinal degeneration
Abstract
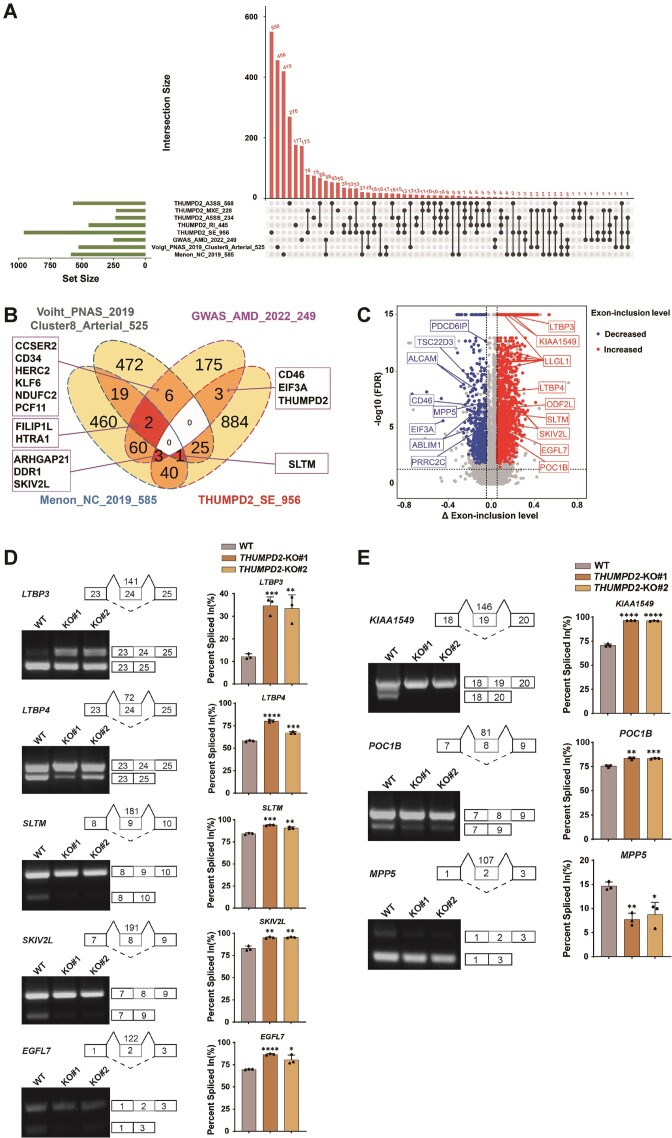
The mechanisms by which the relatively conserved spliceosome manages the enormously large number of splicing events that occur in humans (∼200 000 versus ∼300 in yeast) are poorly understood. Here, we show deposition of one RNA modification-N2-methylguanosine (m2G) on the G72 of U6 snRNA (the catalytic center of the spliceosome) promotes efficient pre-mRNA splicing activity in human cells. This modification was identified to be conserved among vertebrates. Further, THUMPD2 was demonstrated as the methyltransferase responsible for U6 m2G72 by explicitly recognizing the U6-specific sequences and structural elements. The knock-out of THUMPD2 eliminated U6 m2G72 and impaired the pre-mRNA splicing activity, resulting in thousands of changed alternative splicing events of endogenous pre-mRNAs in human cells. Notably, the aberrantly spliced pre-mRNA population elicited the nonsense-mediated mRNA decay pathway. We further show that THUMPD2 was associated with age-related macular degeneration and retinal function. Our study thus demonstrates how an RNA epigenetic modification of the major spliceosome regulates global pre-mRNA splicing and impacts physiology and disease.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures







Similar articles
-
N 2-methylguanosine modifications on human tRNAs and snRNA U6 are important for cell proliferation, protein translation and pre-mRNA splicing.Nucleic Acids Res. 2023 Aug 11;51(14):7496-7519. doi: 10.1093/nar/gkad487. Nucleic Acids Res. 2023. PMID: 37283053 Free PMC article.
-
How Is Precursor Messenger RNA Spliced by the Spliceosome?Annu Rev Biochem. 2020 Jun 20;89:333-358. doi: 10.1146/annurev-biochem-013118-111024. Epub 2019 Dec 9. Annu Rev Biochem. 2020. PMID: 31815536 Review.
-
m6A modification of U6 snRNA modulates usage of two major classes of pre-mRNA 5' splice site.Elife. 2022 Nov 21;11:e78808. doi: 10.7554/eLife.78808. Elife. 2022. PMID: 36409063 Free PMC article.
-
The conserved central domain of yeast U6 snRNA: importance of U2-U6 helix Ia in spliceosome assembly.RNA. 2002 Aug;8(8):997-1010. doi: 10.1017/s1355838202025013. RNA. 2002. PMID: 12212854 Free PMC article.
-
Structural features of U6 snRNA and dynamic interactions with other spliceosomal components leading to pre-mRNA splicing.Biochimie. 1996;78(6):436-42. doi: 10.1016/0300-9084(96)84750-x. Biochimie. 1996. PMID: 8915533 Review.
Cited by
-
THUMPD3 regulates alternative splicing of ECM transcripts in human lung cancer cells and promotes proliferation and migration.PLoS One. 2024 Dec 10;19(12):e0314655. doi: 10.1371/journal.pone.0314655. eCollection 2024. PLoS One. 2024. PMID: 39656728 Free PMC article.
-
RNA Polymerase III-Transcribed RNAs in Health and Disease: Mechanisms, Dysfunction, and Future Directions.Int J Mol Sci. 2025 Jun 18;26(12):5852. doi: 10.3390/ijms26125852. Int J Mol Sci. 2025. PMID: 40565315 Free PMC article. Review.
-
U6 snRNA m6A modification is required for accurate and efficient splicing of C. elegans and human pre-mRNAs.Nucleic Acids Res. 2024 Aug 27;52(15):9139-9160. doi: 10.1093/nar/gkae447. Nucleic Acids Res. 2024. PMID: 38808663 Free PMC article.
References
-
- Brody E., Abelson J.. The “spliceosome”: yeast pre-messenger RNA associates with a 40S complex in a splicing-dependent reaction. Science. 1985; 228:963–967. - PubMed
-
- Grabowski P.J., Seiler S.R., Sharp P.A.. A multicomponent complex is involved in the splicing of messenger RNA precursors. Cell. 1985; 42:345–353. - PubMed
-
- Frendewey D., Keller W.. Stepwise assembly of a pre-mRNA splicing complex requires U-snRNPs and specific intron sequences. Cell. 1985; 42:355–367. - PubMed
-
- Pan Q., Shai O., Lee L.J., Frey B.J., Blencowe B.J.. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008; 40:1413–1415. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
