

Multiscale spatial mapping of cell populations across anatomical sites in healthy human skin and basal cell carcinoma
- PMID: 38165934
- PMCID: PMC10786309
- DOI: 10.1073/pnas.2313326120
Multiscale spatial mapping of cell populations across anatomical sites in healthy human skin and basal cell carcinoma
Abstract
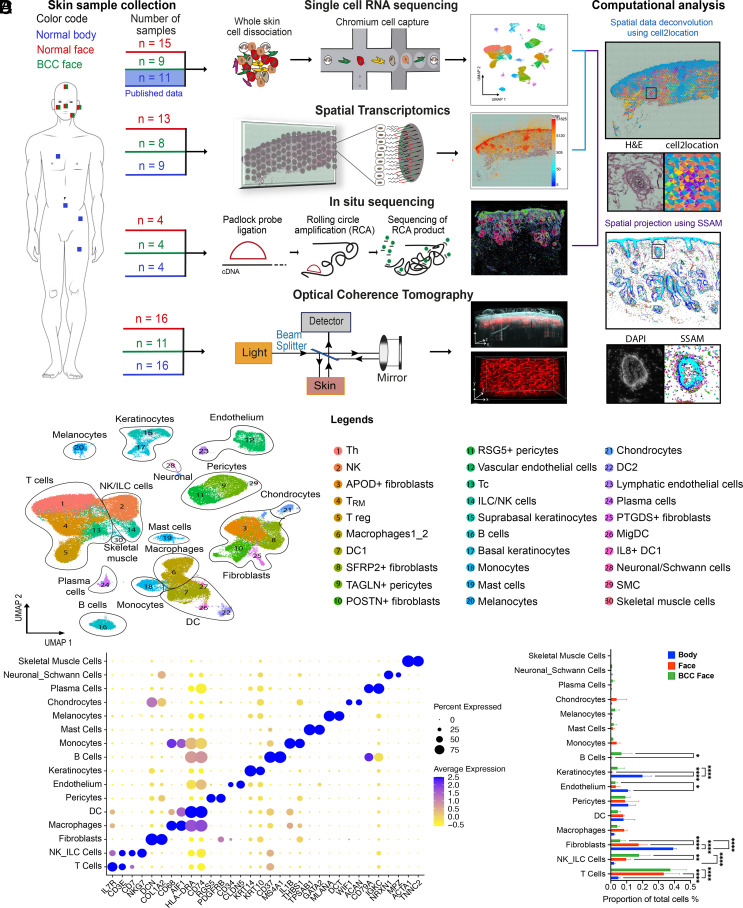
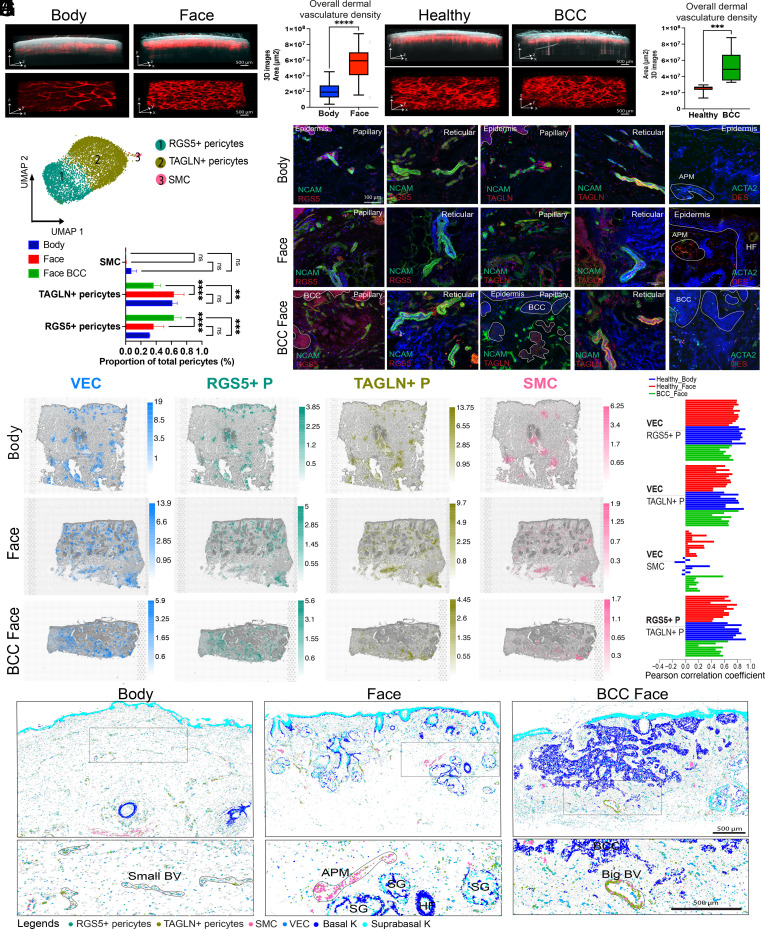
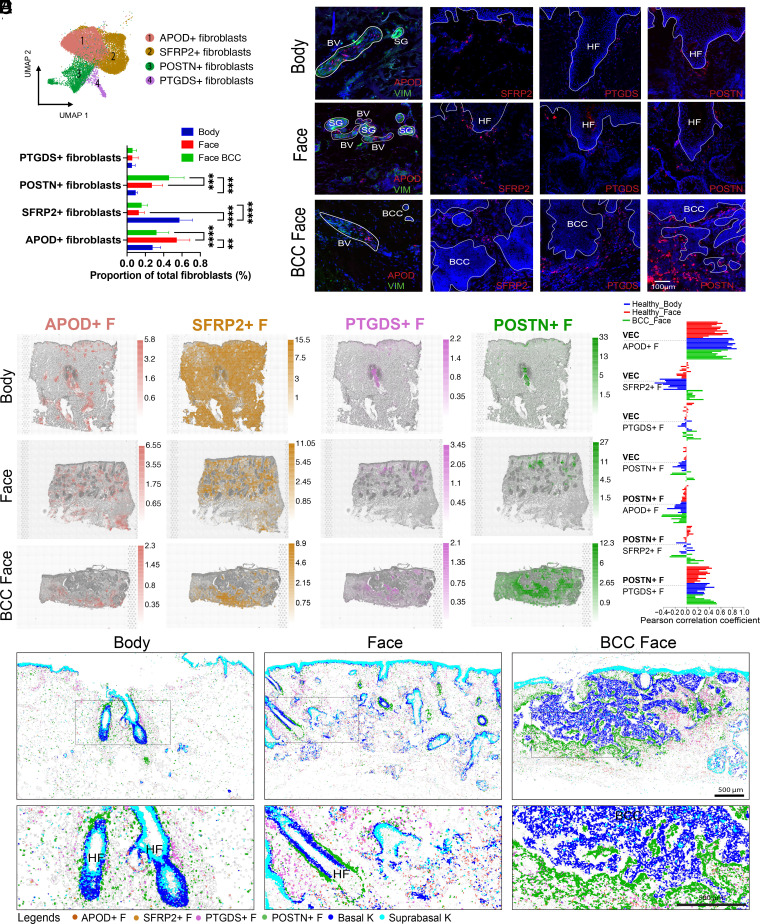
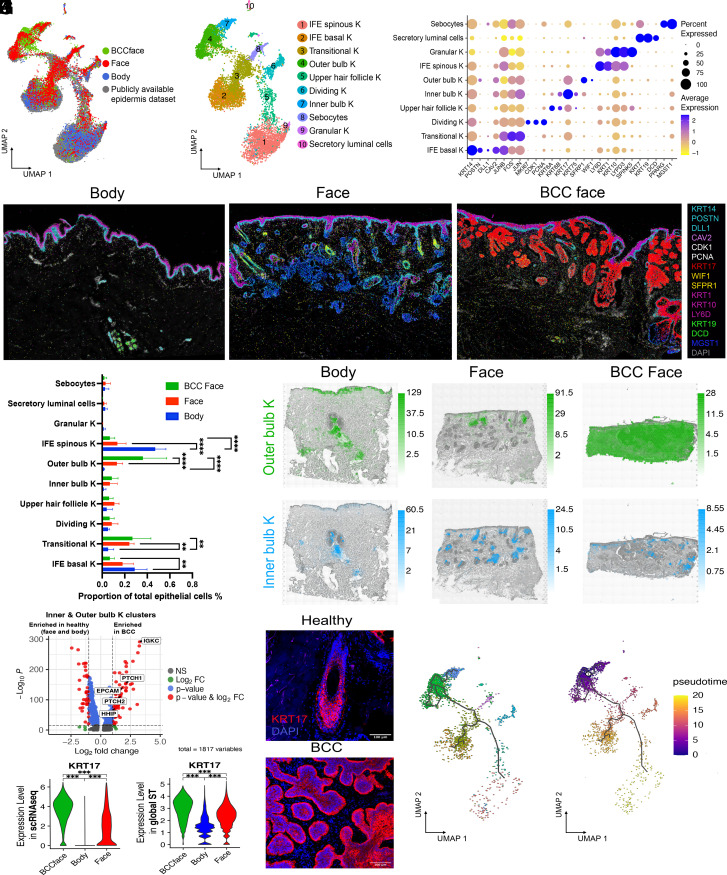
Our understanding of how human skin cells differ according to anatomical site and tumour formation is limited. To address this, we have created a multiscale spatial atlas of healthy skin and basal cell carcinoma (BCC), incorporating in vivo optical coherence tomography, single-cell RNA sequencing, spatial global transcriptional profiling, and in situ sequencing. Computational spatial deconvolution and projection revealed the localisation of distinct cell populations to specific tissue contexts. Although cell populations were conserved between healthy anatomical sites and in BCC, mesenchymal cell populations including fibroblasts and pericytes retained signatures of developmental origin. Spatial profiling and in silico lineage tracing support a hair follicle origin for BCC and demonstrate that cancer-associated fibroblasts are an expansion of a POSTN+ subpopulation associated with hair follicles in healthy skin. RGS5+ pericytes are also expanded in BCC suggesting a role in vascular remodelling. We propose that the identity of mesenchymal cell populations is regulated by signals emanating from adjacent structures and that these signals are repurposed to promote the expansion of skin cancer stroma. The resource we have created is publicly available in an interactive format for the research community.
Keywords: basal cell carcinoma; fibroblasts; human cell atlas; single cell RNA sequencing; skin.
Conflict of interest statement
Competing interests statement:In the last 3 y, S.A.T. has been a remunerated Scientific Advisory Board member for GlaxoSmithKline, Qiagen, Foresite Labs, and is a co-founder and equity holder of TransitionBio. F.M.W. and M.D.L. are co-founders of Fibrodyne. F.M.W. and M.D.L. have filed two patents related to skin fibroblasts.
Figures
References
-
- Crowson A. N., Basal cell carcinoma: Biology, morphology and clinical implications. Mod. Pathol. 19, S127–S147 (2006). - PubMed
-
- Le Lièvre C. S., Le Douarin N. M., Mesenchymal derivatives of the neural crest: Analysis of chimaeric quail and chick embryos. J. Embryol. Exp. Morphol. 34, 125–154 (1975). - PubMed
-
- Driskell R. R., Watt F. M., Understanding fibroblast heterogeneity in the skin. Trends Cell Biol. 25, 92–99 (2015). - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
