

Structural role of K224 in taniborbactam inhibition of NDM-1
- PMID: 38174924
- PMCID: PMC10848753
- DOI: 10.1128/aac.01332-23
Structural role of K224 in taniborbactam inhibition of NDM-1
Abstract
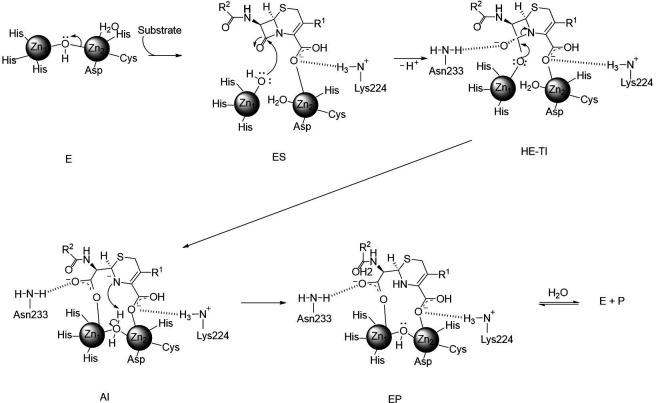
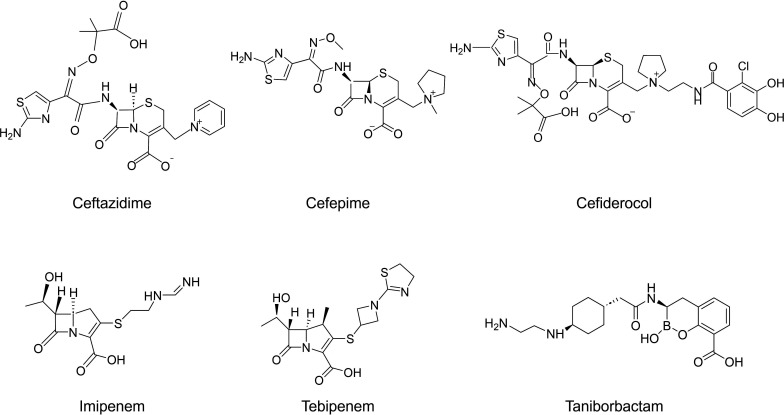
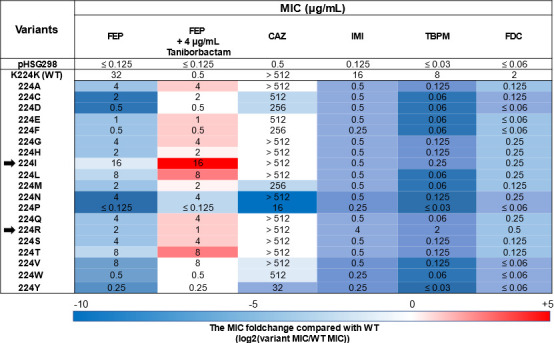
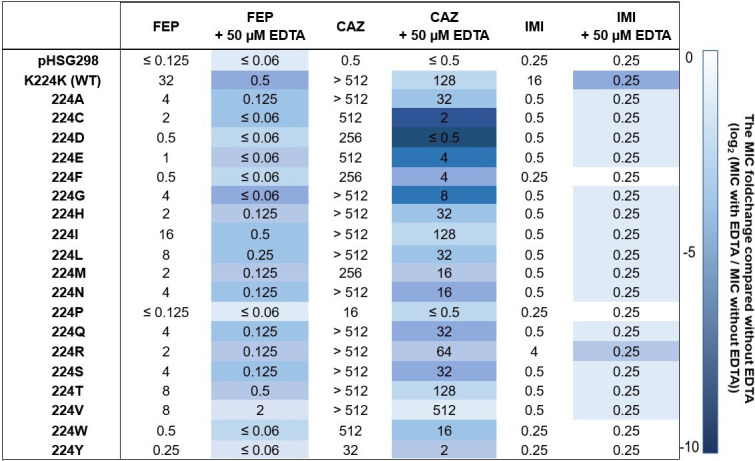
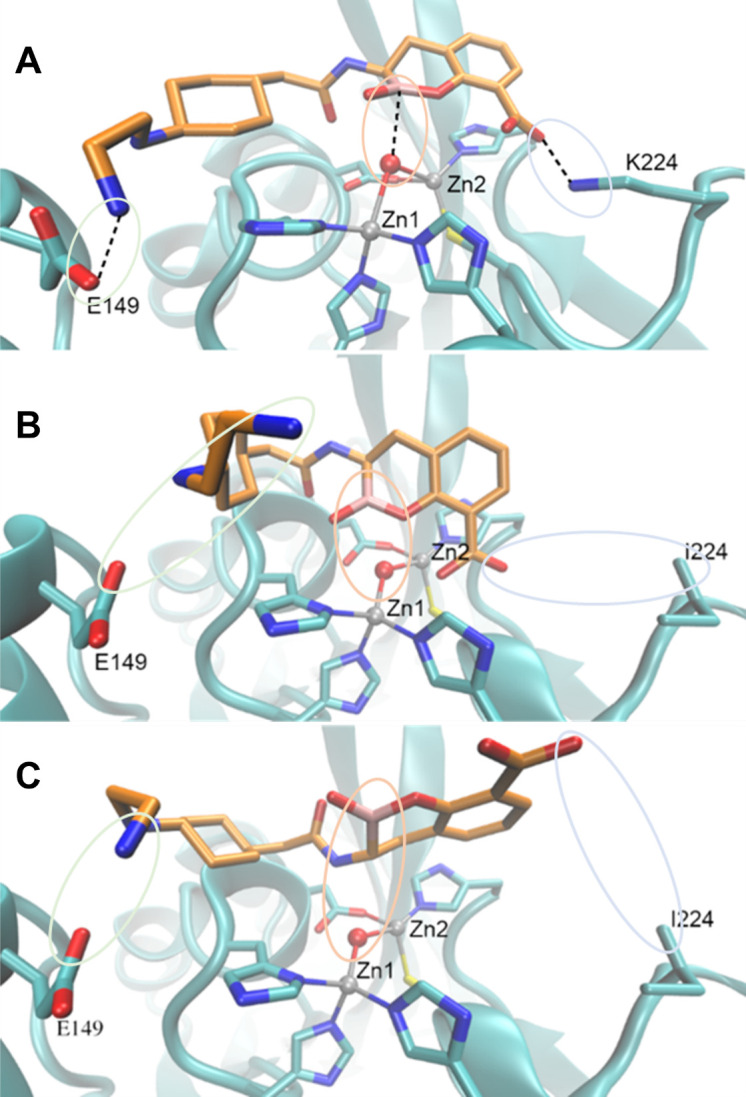
Taniborbactam (TAN; VNRX-5133) is a novel bicyclic boronic acid β-lactamase inhibitor (BLI) being developed in combination with cefepime (FEP). TAN inhibits both serine and some metallo-β-lactamases. Previously, the substitution R228L in VIM-24 was shown to increase activity against oxyimino-cephalosporins like FEP and ceftazidime (CAZ). We hypothesized that substitutions at K224, the homologous position in NDM-1, could impact FEP/TAN resistance. To evaluate this, a library of codon-optimized NDM K224X clones for minimum inhibitory concentration (MIC) measurements was constructed; steady-state kinetics and molecular docking simulations were next performed. Surprisingly, our investigation revealed that the addition of TAN restored FEP susceptibility only for NDM-1, as the MICs for the other 19 K224X variants remained comparable to those of FEP alone. Moreover, compared to NDM-1, all K224X variants displayed significantly lower MICs for imipenem, tebipenem, and cefiderocol (32-, 133-, and 33-fold lower, respectively). In contrast, susceptibility to CAZ was mostly unaffected. Kinetic assays with the K224I variant, the only variant with hydrolytic activity to FEP comparable to NDM-1, confirmed that the inhibitory capacity of TAN was modestly compromised (IC50 0.01 µM vs 0.14 µM for NDM-1). Lastly, structural modeling and docking simulations of TAN in NDM-1 and in the K224I variant revealed that the hydrogen bond between TAN's carboxylate with K224 is essential for the productive binding of TAN to the NDM-1 active site. In addition to the report of NDM-9 (E149K) as FEP/TAN resistant, this study demonstrates the fundamental role of single amino acid substitutions in the inhibition of NDM-1 by TAN.
Keywords: K224; NDM-1; metallo-β-lactamase; taniborbactam; β-lactamase inhibitor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- WHO . 2017. WHO publishes list of bacteria for which new antibiotics are urgently needed. Available from: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-.... Retrieved 10 Oct 2023.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
