

SARM1 regulates NAD+-linked metabolism and select immune genes in macrophages
- PMID: 38176648
- PMCID: PMC10847163
- DOI: 10.1016/j.jbc.2023.105620
SARM1 regulates NAD+-linked metabolism and select immune genes in macrophages
Abstract
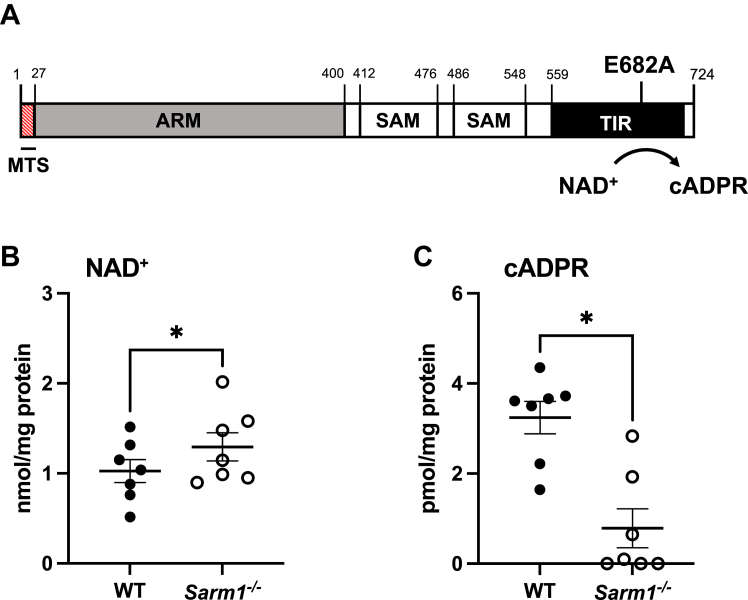
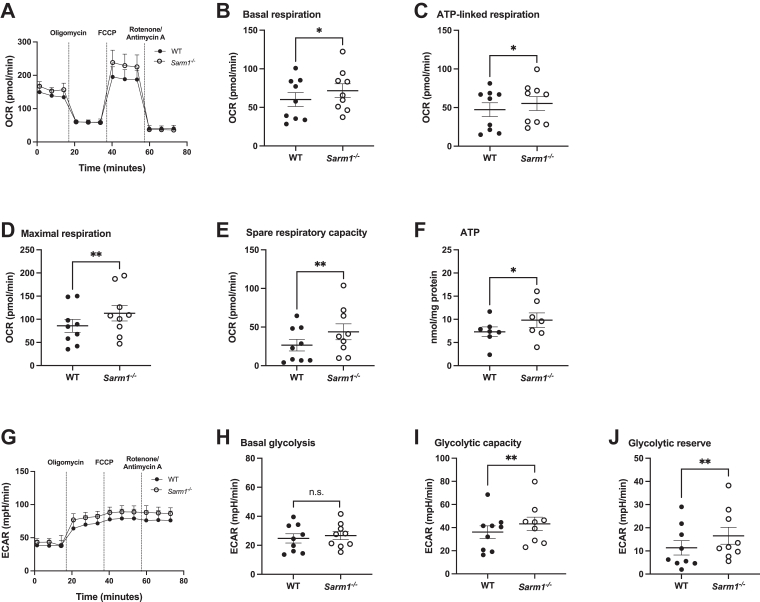
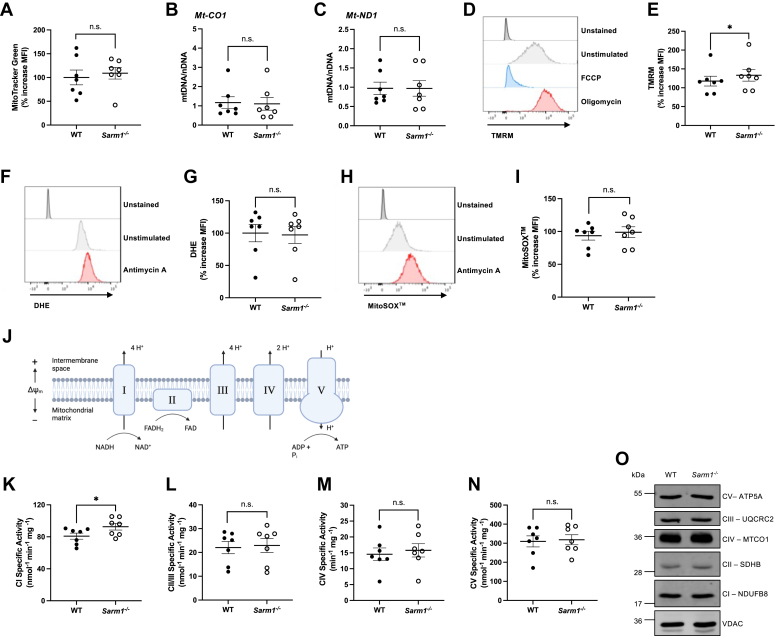
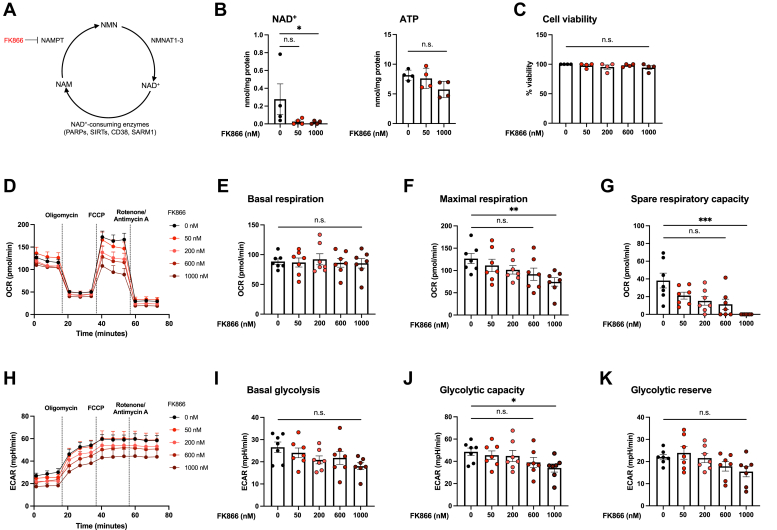
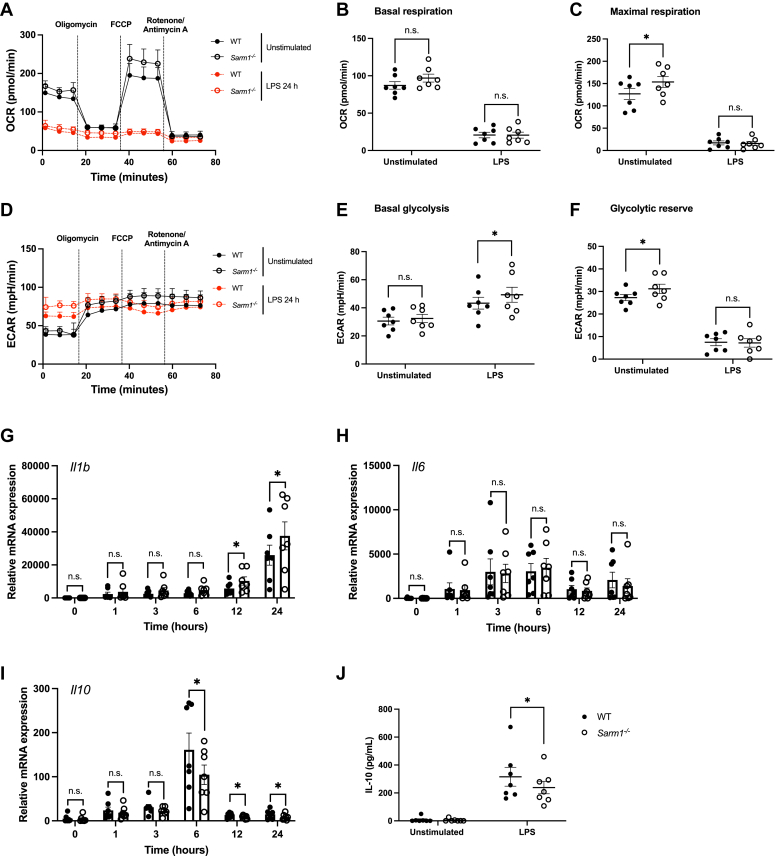
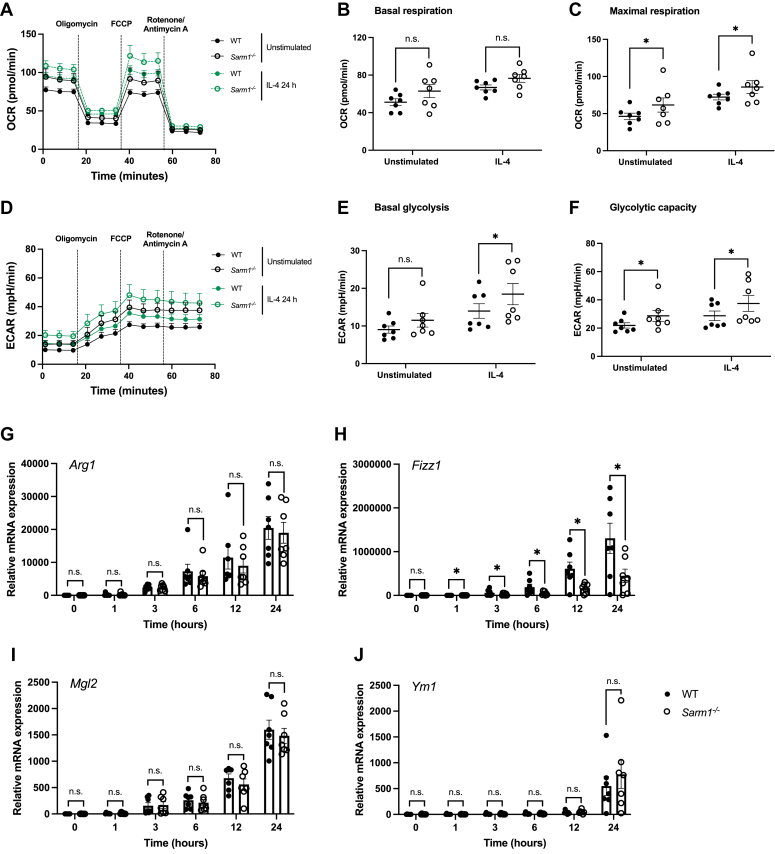
Sterile alpha and HEAT/armadillo motif-containing protein (SARM1) was recently described as a NAD+-consuming enzyme and has previously been shown to regulate immune responses in macrophages. Neuronal SARM1 is known to contribute to axon degeneration due to its NADase activity. However, how SARM1 affects macrophage metabolism has not been explored. Here, we show that macrophages from Sarm1-/- mice display elevated NAD+ concentrations and lower cyclic ADP-ribose, a known product of SARM1-dependent NAD+ catabolism. Further, SARM1-deficient macrophages showed an increase in the reserve capacity of oxidative phosphorylation and glycolysis compared to WT cells. Stimulation of macrophages to a proinflammatory state by lipopolysaccharide (LPS) revealed that SARM1 restricts the ability of macrophages to upregulate glycolysis and limits the expression of the proinflammatory gene interleukin (Il) 1b, but boosts expression of anti-inflammatory Il10. In contrast, we show macrophages lacking SARM1 induced to an anti-inflammatory state by IL-4 stimulation display increased oxidative phosphorylation and glycolysis, and reduced expression of the anti-inflammatory gene, Fizz1. Overall, these data show that SARM1 fine-tunes immune gene transcription in macrophages via consumption of NAD+ and altered macrophage metabolism.
Keywords: NAD(+); NADase; SARM1; cADPR; cytokine induction; macrophages; metabolism.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
