

Intestinal Bacteroides modulates inflammation, systemic cytokines, and microbial ecology via propionate in a mouse model of cystic fibrosis
- PMID: 38179971
- PMCID: PMC10865972
- DOI: 10.1128/mbio.03144-23
Intestinal Bacteroides modulates inflammation, systemic cytokines, and microbial ecology via propionate in a mouse model of cystic fibrosis
Abstract
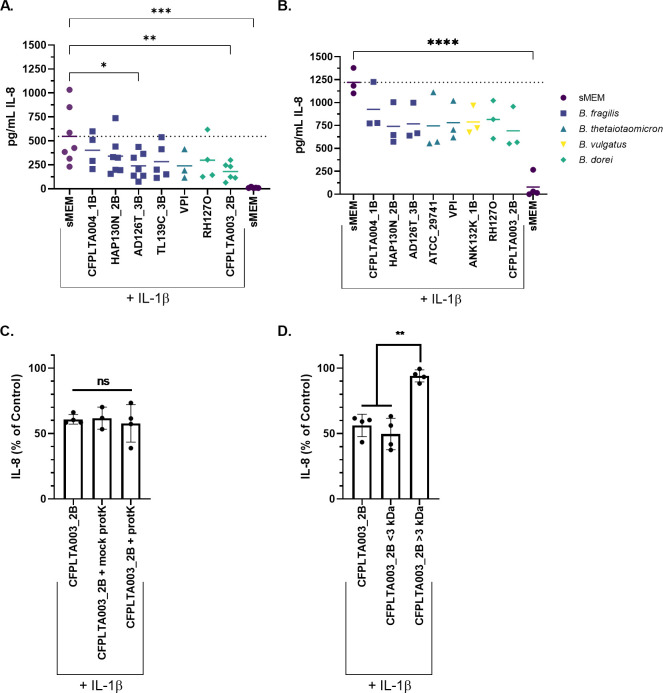
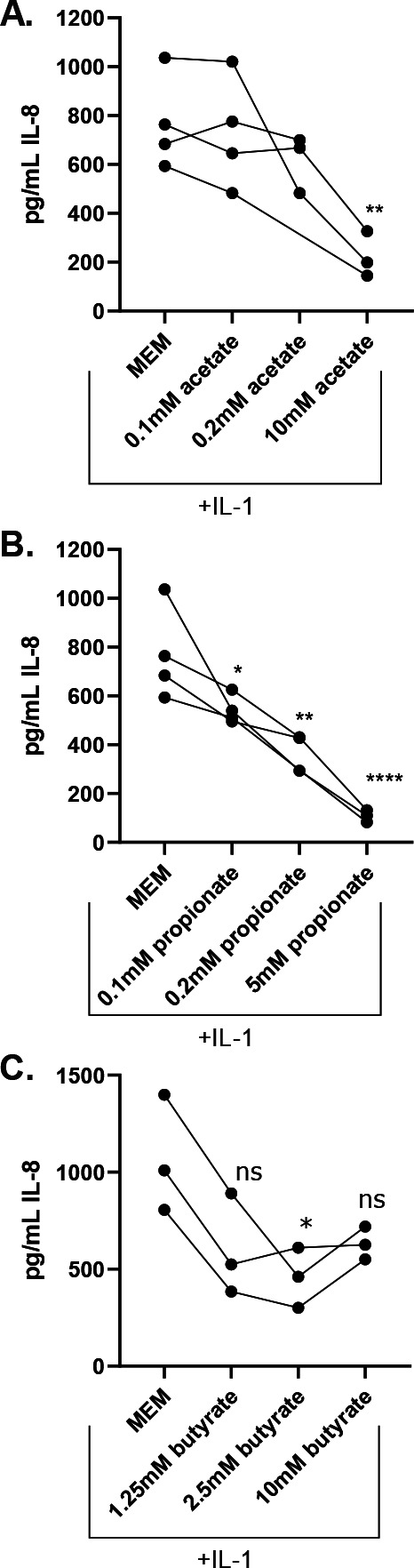
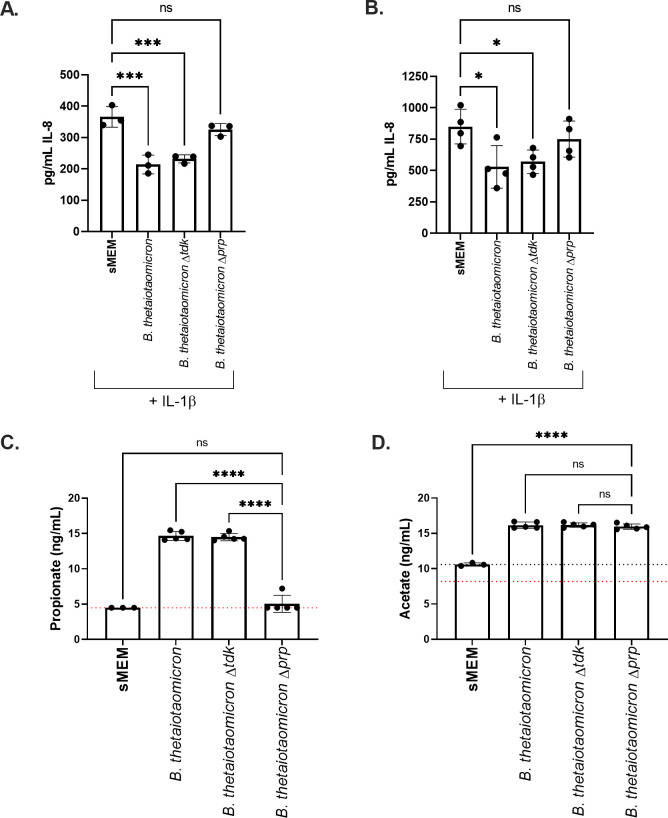
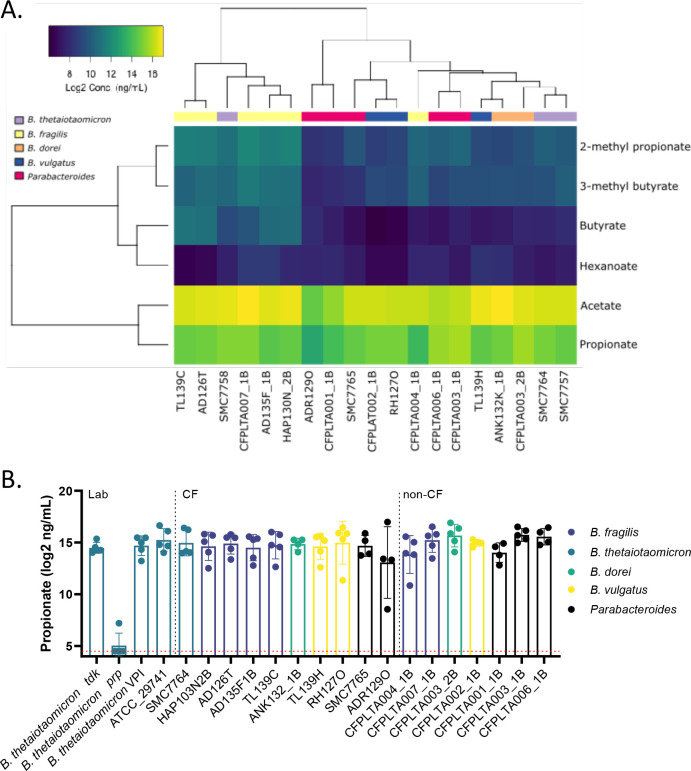
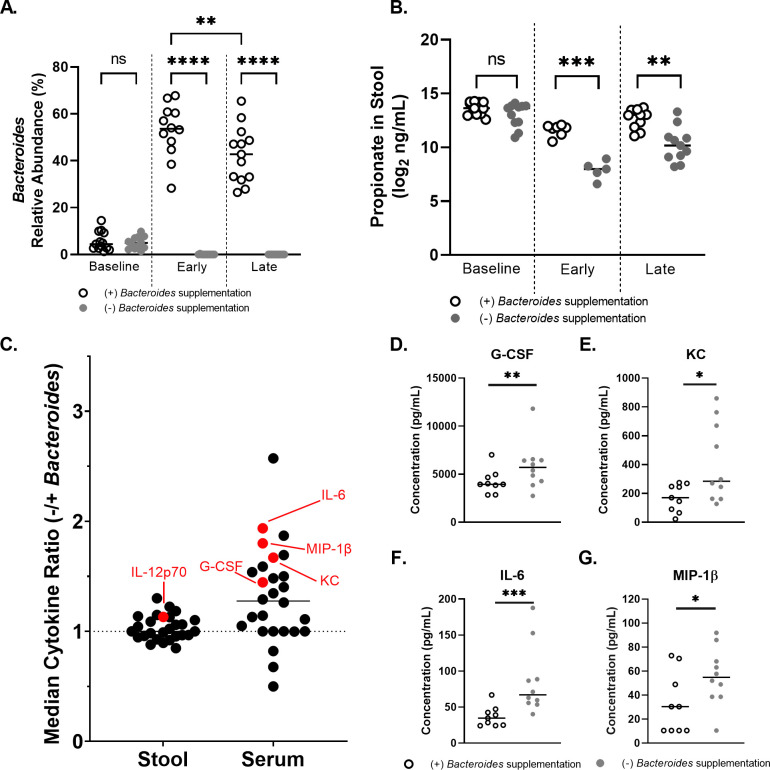
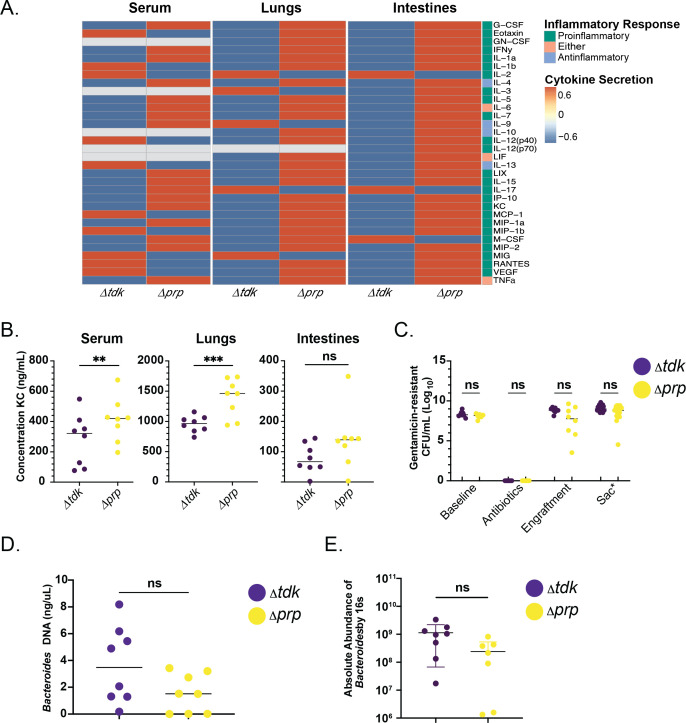
Persons with cystic fibrosis (CF), starting in early life, show intestinal microbiome dysbiosis characterized in part by a decreased relative abundance of the genus Bacteroides. Bacteroides is a major producer of the intestinal short chain fatty acid propionate. We demonstrate here that cystic fibrosis transmembrane conductance regulator-defective (CFTR-/-) Caco-2 intestinal epithelial cells are responsive to the anti-inflammatory effects of propionate. Furthermore, Bacteroides isolates inhibit the IL-1β-induced inflammatory response of CFTR-/- Caco-2 intestinal epithelial cells and do so in a propionate-dependent manner. The introduction of Bacteroides-supplemented stool from infants with cystic fibrosis into the gut of CftrF508del mice results in higher propionate in the stool as well as the reduction in several systemic pro-inflammatory cytokines. Bacteroides supplementation also reduced the fecal relative abundance of Escherichia coli, indicating a potential interaction between these two microbes, consistent with previous clinical studies. For a Bacteroides propionate mutant in the mouse model, pro-inflammatory cytokine KC is higher in the airway and serum compared with the wild-type (WT) strain, with no significant difference in the absolute abundance of these two strains. Taken together, our data indicate the potential multiple roles of Bacteroides-derived propionate in the modulation of systemic and airway inflammation and mediating the intestinal ecology of infants and children with CF. The roles of Bacteroides and the propionate it produces may help explain the observed gut-lung axis in CF and could guide the development of probiotics to mitigate systemic and airway inflammation for persons with CF.IMPORTANCEThe composition of the gut microbiome in persons with CF is correlated with lung health outcomes, a phenomenon referred to as the gut-lung axis. Here, we demonstrate that the intestinal microbe Bacteroides decreases inflammation through the production of the short-chain fatty acid propionate. Supplementing the levels of Bacteroides in an animal model of CF is associated with reduced systemic inflammation and reduction in the relative abundance of the opportunistically pathogenic group Escherichia/Shigella in the gut. Taken together, these data demonstrate a key role for Bacteroides and microbially produced propionate in modulating inflammation, gut microbial ecology, and the gut-lung axis in cystic fibrosis. These data support the role of Bacteroides as a potential probiotic in CF.
Keywords: SCFA; cystic fibrosis; gut; inflammation; probiotic; propionate.
Conflict of interest statement
We have filed and have been issued a patent relevant to this work: U.S. Provisional Application No. 16/979,824, filed 28 March 2018, issued 27 June 2022: Altering the Intestinal Microbiome In Cystic Fibrosis; inventors: J. C. Madan and G. A. O'Toole.
Figures
References
-
- Registry C. 2019. Annual data report. Bethesda, Maryland 2020 Cystic Fibrosis Foundation
-
- Riordan JR, Rommens JM, Kerem B-S, Alon N, Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou J-L, Drumm ML, Iannuzzi MC, Collins FS, Tsui L-C. 1989. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245:1066–1073. doi: 10.1126/science.2475911 - DOI - PubMed
-
- Antosca KM, Chernikova DA, Price CE, Ruoff KL, Li K, Guill MF, Sontag NR, Morrison HG, Hao S, Drumm ML, MacKenzie TA, Dorman DB, Feenan LM, Williams MA, Dessaint J, Yuan IH, Aldrich BJ, Moulton LA, Ting L, Martinez-Del Campo A, Stewart EJ, Karagas MR, O’Toole GA, Madan JC. 2019. Altered stool microbiota of infants with cystic fibrosis shows a reduction in genera associated with immune programming from birth. J Bacteriol 201:e00274-19. doi: 10.1128/JB.00274-19 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
