

USP1-dependent nucleolytic expansion of PRIMPOL-generated nascent DNA strand discontinuities during replication stress
- PMID: 38180818
- PMCID: PMC10954467
- DOI: 10.1093/nar/gkad1237
USP1-dependent nucleolytic expansion of PRIMPOL-generated nascent DNA strand discontinuities during replication stress
Abstract
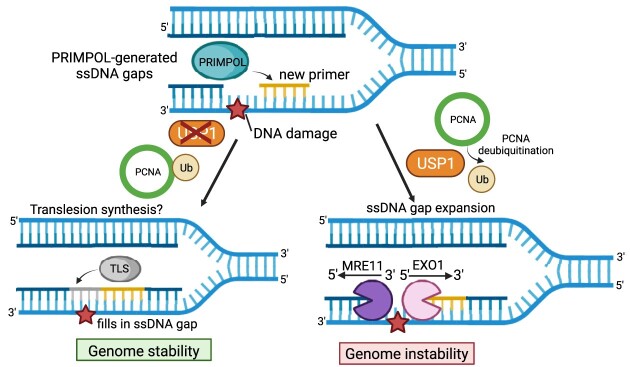
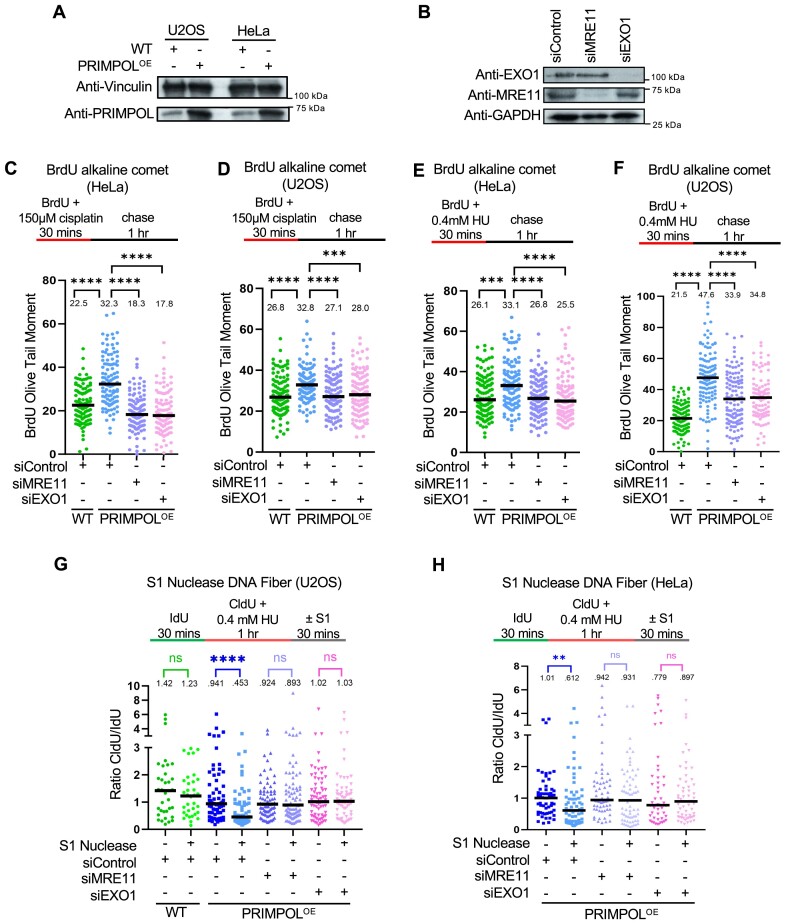
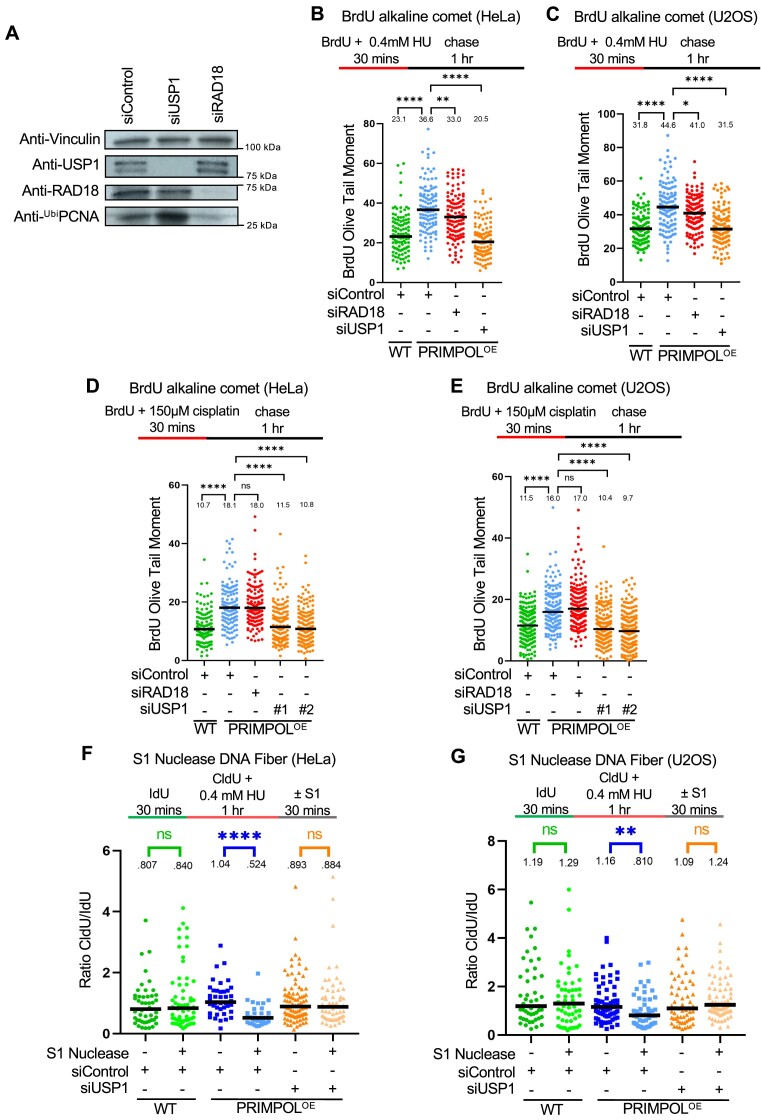
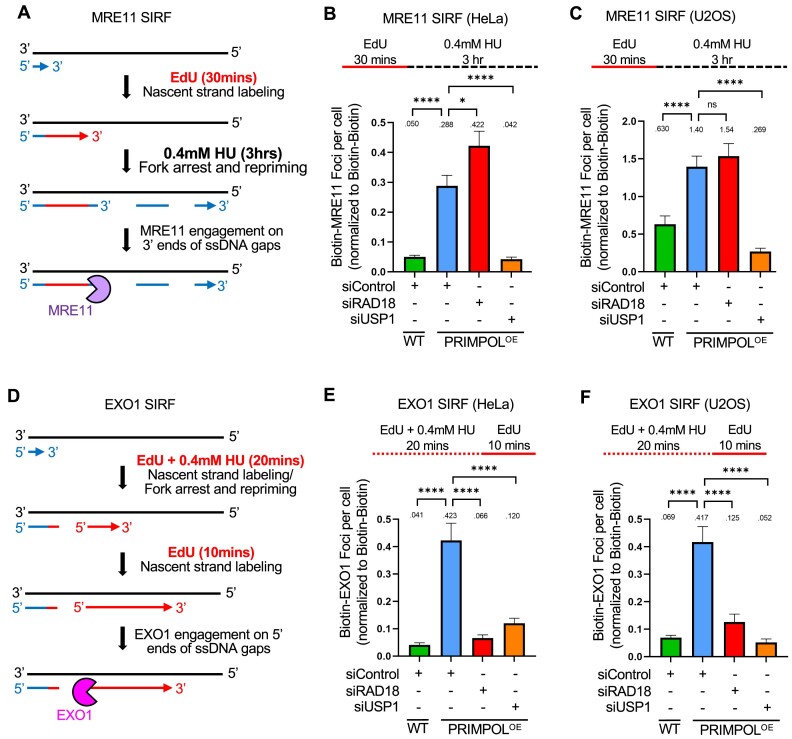
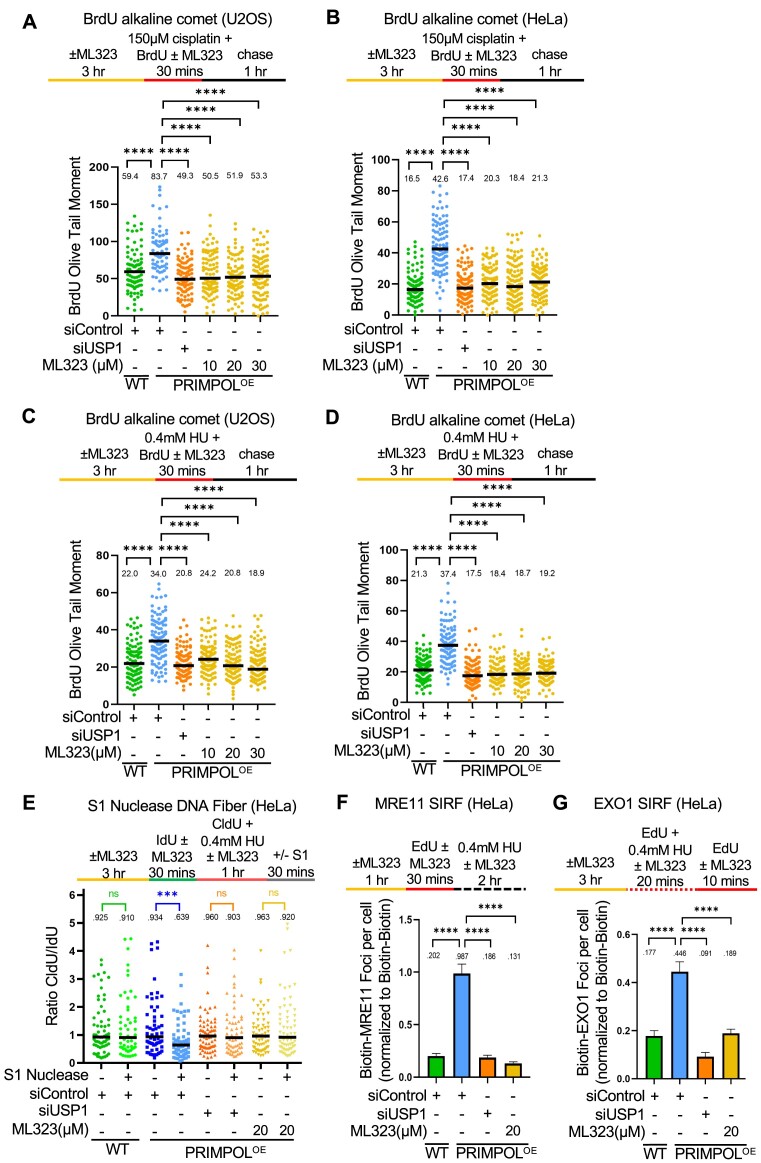
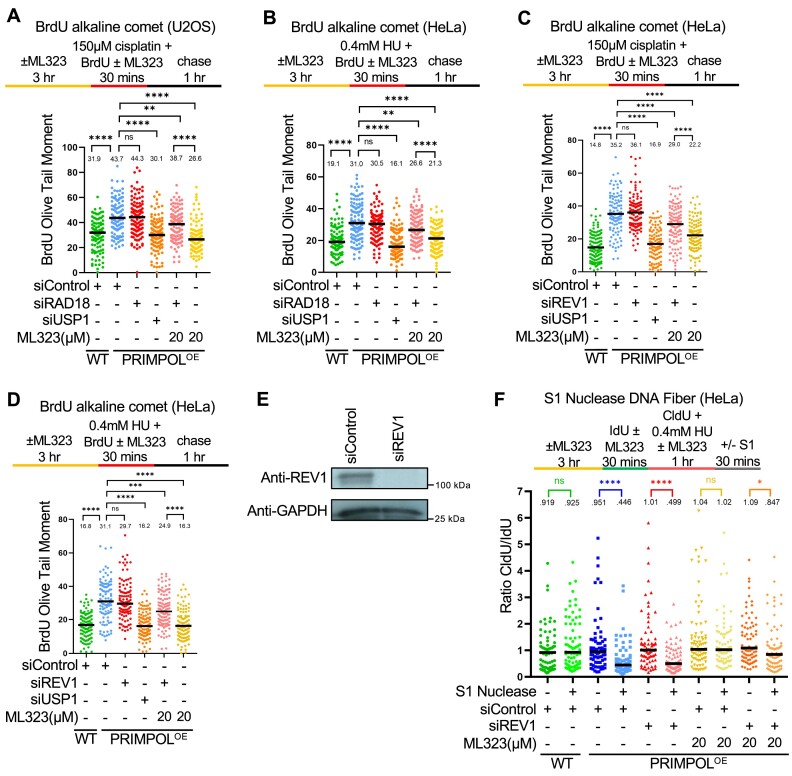
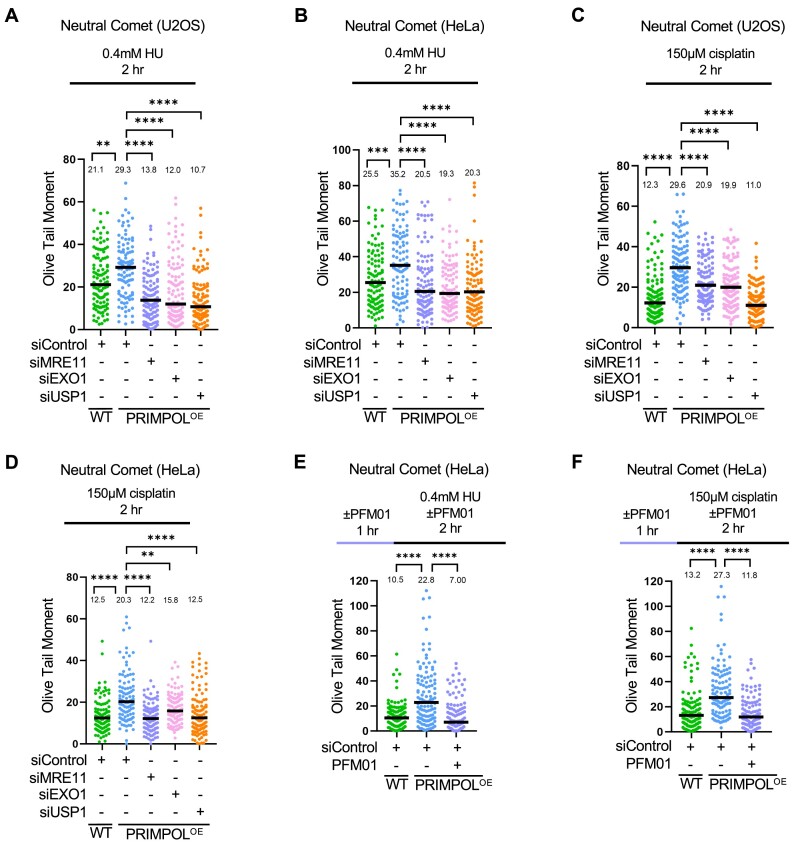
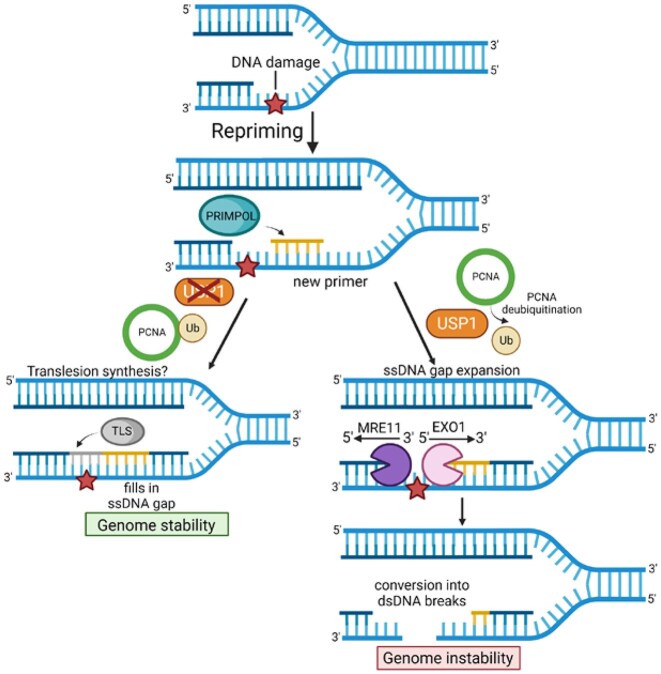
DNA replication stress-induced fork arrest represents a significant threat to genomic integrity. One major mechanism of replication restart involves repriming downstream of the arrested fork by PRIMPOL, leaving behind a single-stranded DNA (ssDNA) gap. Accumulation of nascent strand ssDNA gaps has emerged as a possible determinant of the cellular hypersensitivity to genotoxic agents in certain genetic backgrounds such as BRCA deficiency, but how gaps are converted into cytotoxic structures is still unclear. Here, we investigate the processing of PRIMPOL-dependent ssDNA gaps upon replication stress induced by hydroxyurea and cisplatin. We show that gaps generated in PRIMPOL-overexpressing cells are expanded in the 3'-5' direction by the MRE11 exonuclease, and in the 5'-3' direction by the EXO1 exonuclease. This bidirectional exonucleolytic gap expansion ultimately promotes their conversion into DSBs. We moreover identify the de-ubiquitinating enzyme USP1 as a critical regulator of PRIMPOL-generated ssDNA gaps. USP1 promotes gap accumulation during S-phase, and their expansion by the MRE11 and EXO1 nucleases. This activity of USP1 is linked to its role in de-ubiquitinating PCNA, suggesting that PCNA ubiquitination prevents gap accumulation during replication. Finally, we show that USP1 depletion suppresses DSB formation in PRIMPOL-overexpressing cells, highlighting an unexpected role for USP1 in promoting genomic instability under these conditions.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Temporally distinct post-replicative repair mechanisms fill PRIMPOL-dependent ssDNA gaps in human cells.Mol Cell. 2021 Oct 7;81(19):4026-4040.e8. doi: 10.1016/j.molcel.2021.09.013. Mol Cell. 2021. PMID: 34624216 Free PMC article.
-
Multi-step processing of replication stress-derived nascent strand DNA gaps by MRE11 and EXO1 nucleases.Nat Commun. 2023 Oct 7;14(1):6265. doi: 10.1038/s41467-023-42011-0. Nat Commun. 2023. PMID: 37805499 Free PMC article.
-
PRIMPOL-Mediated Adaptive Response Suppresses Replication Fork Reversal in BRCA-Deficient Cells.Mol Cell. 2020 Feb 6;77(3):461-474.e9. doi: 10.1016/j.molcel.2019.10.008. Epub 2019 Oct 29. Mol Cell. 2020. PMID: 31676232 Free PMC article.
-
Tolerating DNA damage by repriming: Gap filling in the spotlight.DNA Repair (Amst). 2024 Oct;142:103758. doi: 10.1016/j.dnarep.2024.103758. Epub 2024 Aug 30. DNA Repair (Amst). 2024. PMID: 39236419 Review.
-
Role of Translesion DNA Synthesis in the Metabolism of Replication-associated Nascent Strand Gaps.J Mol Biol. 2024 Jan 1;436(1):168275. doi: 10.1016/j.jmb.2023.168275. Epub 2023 Sep 13. J Mol Biol. 2024. PMID: 37714300 Free PMC article. Review.
Cited by
-
CRISPR knockout genome-wide screens identify the HELQ-RAD52 axis in regulating the repair of cisplatin-induced single-stranded DNA gaps.Nucleic Acids Res. 2024 Dec 11;52(22):13832-13848. doi: 10.1093/nar/gkae998. Nucleic Acids Res. 2024. PMID: 39530221 Free PMC article.
-
CAF-1 promotes efficient PrimPol recruitment to nascent DNA for single-stranded DNA gap formation.Nucleic Acids Res. 2024 Dec 11;52(22):13865-13880. doi: 10.1093/nar/gkae1068. Nucleic Acids Res. 2024. PMID: 39558157 Free PMC article.
-
USP1/UAF1 targets polyubiquitinated PCNA with an exo-cleavage mechanism that can temporarily enrich for monoubiquitinated PCNA.Nat Commun. 2025 Jul 30;16(1):6991. doi: 10.1038/s41467-025-61768-0. Nat Commun. 2025. PMID: 40739138 Free PMC article.
-
Single-Stranded DNA Gap Accumulation Is a Functional Biomarker for USP1 Inhibitor Sensitivity.Cancer Res. 2024 Oct 15;84(20):3435-3446. doi: 10.1158/0008-5472.CAN-23-4007. Cancer Res. 2024. PMID: 38885312 Free PMC article.
-
PARP10 promotes the repair of nascent strand DNA gaps through RAD18 mediated translesion synthesis.Nat Commun. 2024 Jul 23;15(1):6197. doi: 10.1038/s41467-024-50429-3. Nat Commun. 2024. PMID: 39043663 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
