

Interaction molecular QTL mapping discovers cellular and environmental modifiers of genetic regulatory effects
- PMID: 38181730
- PMCID: PMC10806864
- DOI: 10.1016/j.ajhg.2023.11.013
Interaction molecular QTL mapping discovers cellular and environmental modifiers of genetic regulatory effects
Abstract
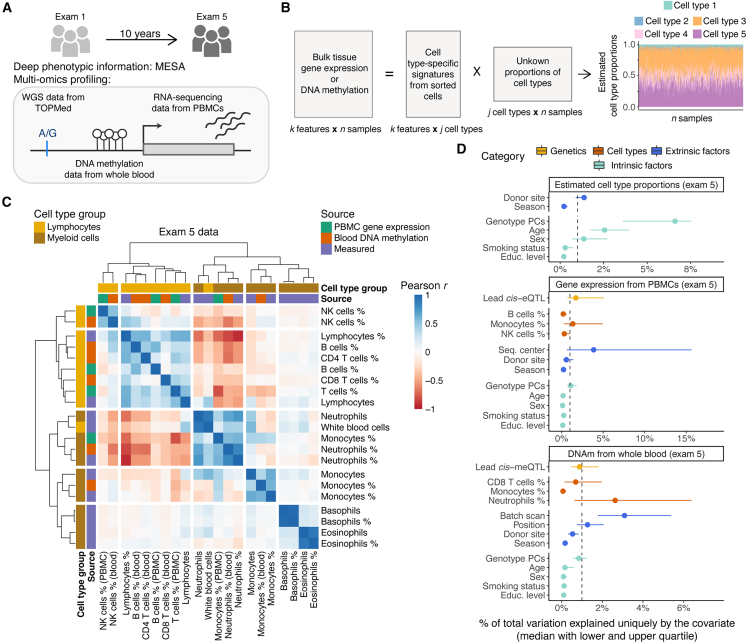
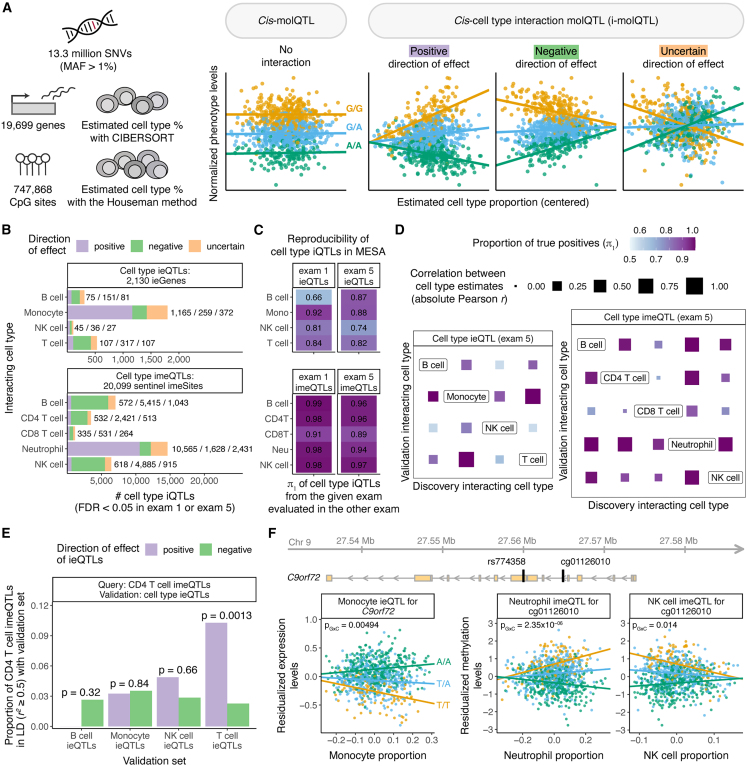
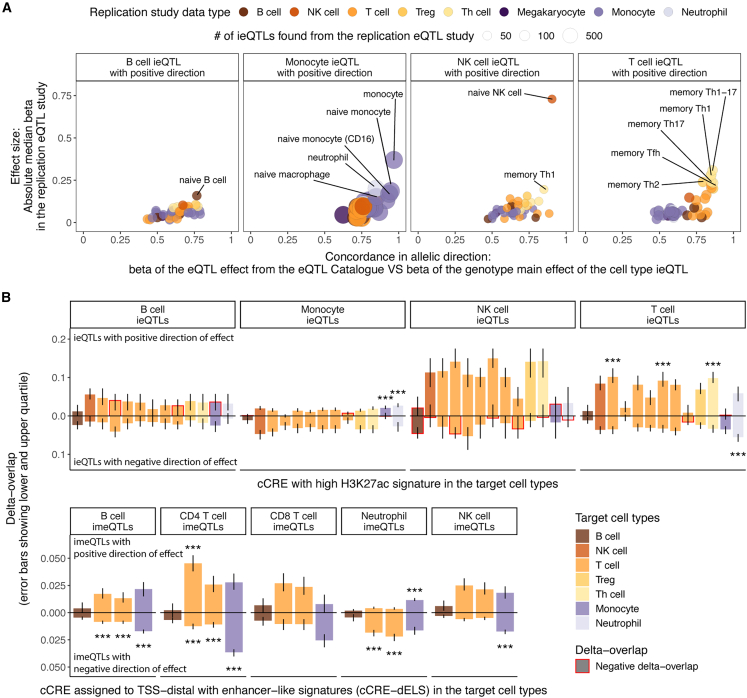
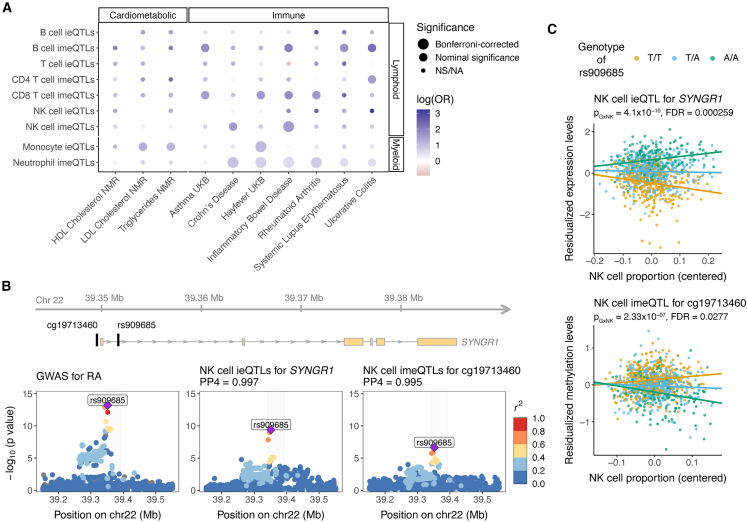
Bulk-tissue molecular quantitative trait loci (QTLs) have been the starting point for interpreting disease-associated variants, and context-specific QTLs show particular relevance for disease. Here, we present the results of mapping interaction QTLs (iQTLs) for cell type, age, and other phenotypic variables in multi-omic, longitudinal data from the blood of individuals of diverse ancestries. By modeling the interaction between genotype and estimated cell-type proportions, we demonstrate that cell-type iQTLs could be considered as proxies for cell-type-specific QTL effects, particularly for the most abundant cell type in the tissue. The interpretation of age iQTLs, however, warrants caution because the moderation effect of age on the genotype and molecular phenotype association could be mediated by changes in cell-type composition. Finally, we show that cell-type iQTLs contribute to cell-type-specific enrichment of diseases that, in combination with additional functional data, could guide future functional studies. Overall, this study highlights the use of iQTLs to gain insights into the context specificity of regulatory effects.
Keywords: DNA methylation; cell-type composition; gene expression; gene-environment interaction; interaction QTL.
Copyright © 2023 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests F.A. is an employee of Illumina, Inc. and an inventor on a patent application related to TensorQTL. T.L. advises Variant Bio, Goldfinch Bio, GlaxoSmithKline, and Pfizer and has equity in Variant Bio.
Figures

Update of
-
Interaction molecular QTL mapping discovers cellular and environmental modifiers of genetic regulatory effects.bioRxiv [Preprint]. 2023 Jun 29:2023.06.26.546528. doi: 10.1101/2023.06.26.546528. bioRxiv. 2023. Update in: Am J Hum Genet. 2024 Jan 4;111(1):133-149. doi: 10.1016/j.ajhg.2023.11.013. PMID: 37425716 Free PMC article. Updated. Preprint.
References
-
- Võsa U., Claringbould A., Westra H.-J., Bonder M.J., Deelen P., Zeng B., Kirsten H., Saha A., Kreuzhuber R., Yazar S., et al. Large-scale cis- and trans-eQTL analyses identify thousands of genetic loci and polygenic scores that regulate blood gene expression. Nat. Genet. 2021;53:1300–1310. - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
