

Enhanced protein secretion in reduced genome strains of Streptomyces lividans
- PMID: 38183102
- PMCID: PMC10768272
- DOI: 10.1186/s12934-023-02269-x
Enhanced protein secretion in reduced genome strains of Streptomyces lividans
Abstract
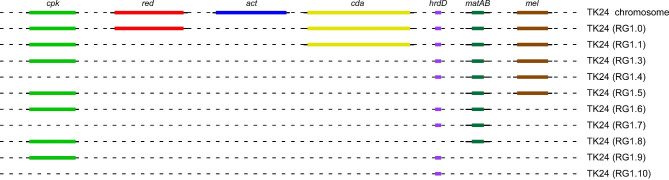
Background: S. lividans TK24 is a popular host for the production of small molecules and the secretion of heterologous protein. Within its large genome, twenty-nine non-essential clusters direct the biosynthesis of secondary metabolites. We had previously constructed ten chassis strains, carrying deletions in various combinations of specialized metabolites biosynthetic clusters, such as those of the blue actinorhodin (act), the calcium-dependent antibiotic (cda), the undecylprodigiosin (red), the coelimycin A (cpk) and the melanin (mel) clusters, as well as the genes hrdD, encoding a non-essential sigma factor, and matAB, a locus affecting mycelial aggregation. Genome reduction was aimed at reducing carbon flow toward specialized metabolite biosynthesis to optimize the production of secreted heterologous protein.
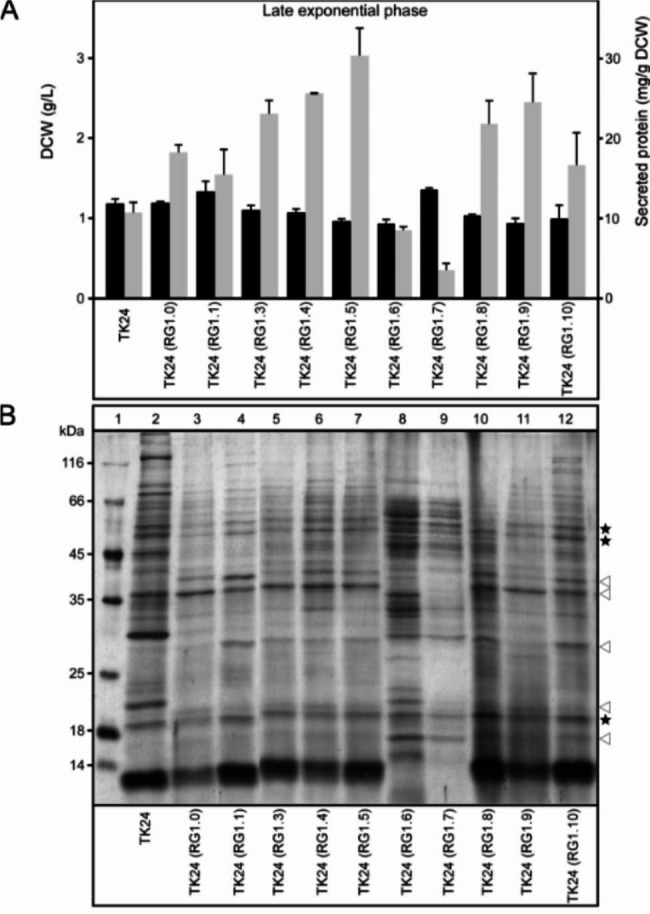
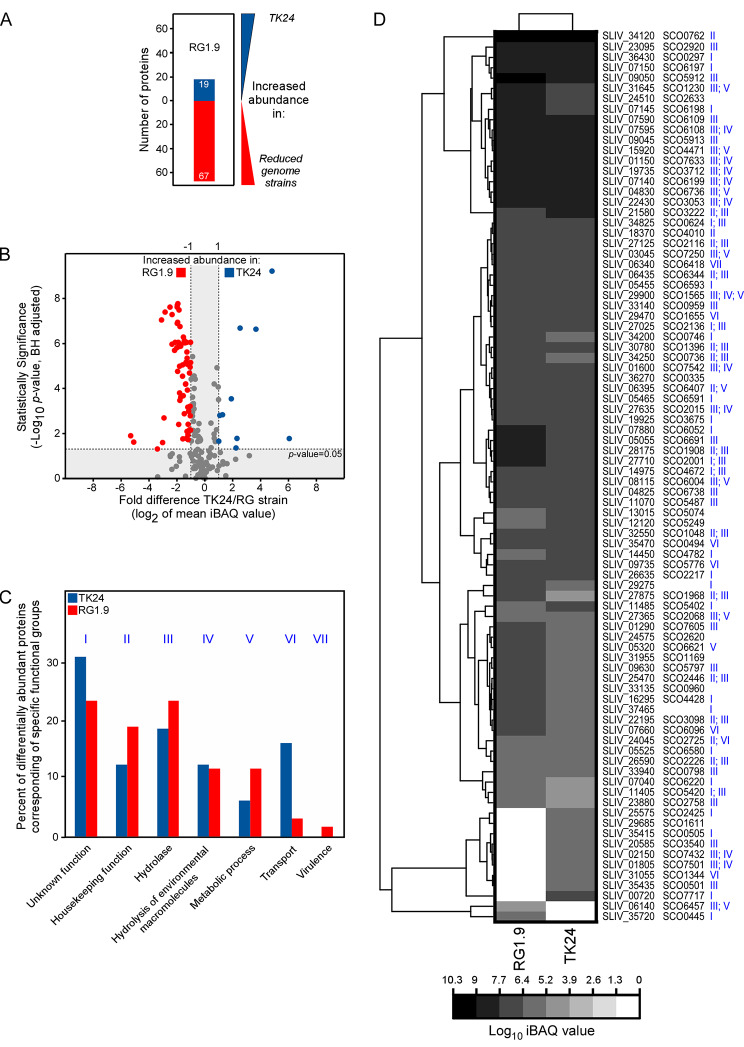
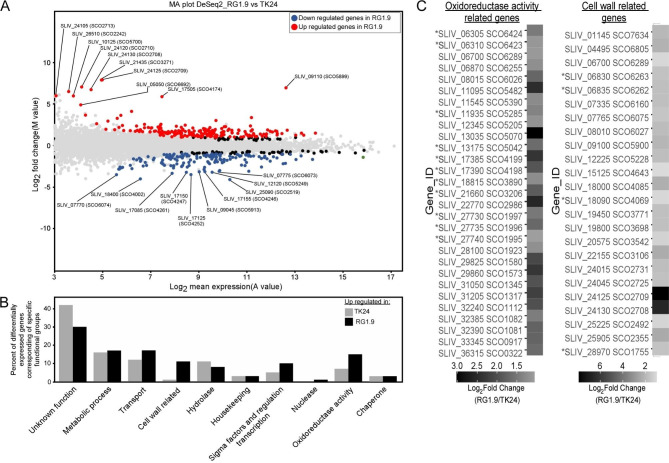
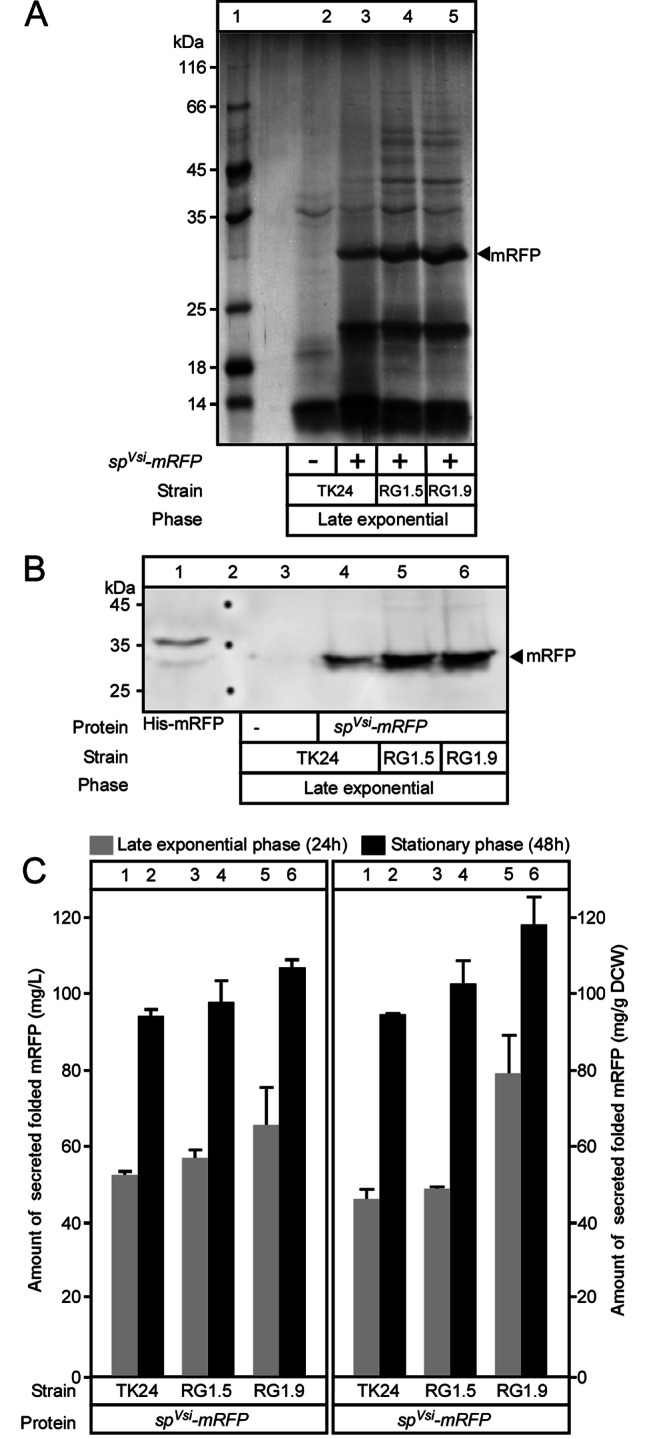
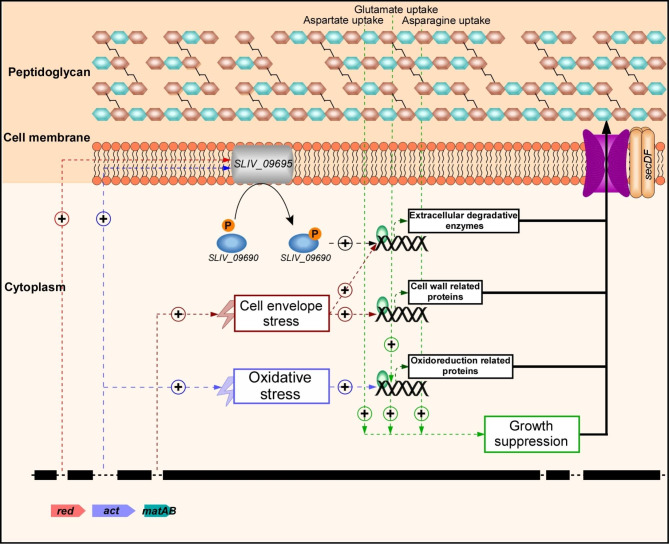
Results: Two of these S. lividans TK24 derived chassis strains showed ~ 15% reduction in biomass yield, 2-fold increase of their total native secretome mass yield and enhanced abundance of several secreted proteins compared to the parental strain. RNAseq and proteomic analysis of the secretome suggested that genome reduction led to cell wall and oxidative stresses and was accompanied by the up-regulation of secretory chaperones and of secDF, a Sec-pathway component. Interestingly, the amount of the secreted heterologous proteins mRFP and mTNFα, by one of these strains, was 12 and 70% higher, respectively, than that secreted by the parental strain.
Conclusion: The current study described a strategy to construct chassis strains with enhanced secretory abilities and proposed a model linking the deletion of specialized metabolite biosynthetic clusters to improved production of secreted heterologous proteins.
Keywords: Heterologous secretion; Proteomics; Reduced genome strains; Secretome; Transcriptomics.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Engineering of Streptomyces lividans for heterologous expression of secondary metabolite gene clusters.Microb Cell Fact. 2020 Jan 9;19(1):5. doi: 10.1186/s12934-020-1277-8. Microb Cell Fact. 2020. PMID: 31918711 Free PMC article.
-
Effects of Increased NADPH Concentration by Metabolic Engineering of the Pentose Phosphate Pathway on Antibiotic Production and Sporulation in Streptomyces lividans TK24.J Microbiol Biotechnol. 2017 Oct 28;27(10):1867-1876. doi: 10.4014/jmb.1707.07046. J Microbiol Biotechnol. 2017. PMID: 28838222
-
An efficient system for stable markerless integration of large biosynthetic gene clusters into Streptomyces chromosomes.Appl Microbiol Biotechnol. 2021 Mar;105(5):2123-2137. doi: 10.1007/s00253-021-11161-w. Epub 2021 Feb 10. Appl Microbiol Biotechnol. 2021. PMID: 33564923
-
Streptomyces protein secretion and its application in biotechnology.FEMS Microbiol Lett. 2018 Nov 1;365(22). doi: 10.1093/femsle/fny250. FEMS Microbiol Lett. 2018. PMID: 30299471 Review.
-
Streptomyces: host for refactoring of diverse bioactive secondary metabolites.3 Biotech. 2021 Jul;11(7):340. doi: 10.1007/s13205-021-02872-y. Epub 2021 Jun 16. 3 Biotech. 2021. PMID: 34221811 Free PMC article. Review.
Cited by
-
Combining culture optimization and synthetic biology to improve production and detection of secondary metabolites in Myxococcus xanthus: application to myxoprincomide.Microbiol Spectr. 2024 Oct 21;12(12):e0174024. doi: 10.1128/spectrum.01740-24. Online ahead of print. Microbiol Spectr. 2024. PMID: 39431896 Free PMC article.
References
-
- Tarkka M, Hampp R. Secondary Metabolites of Soil Streptomycetes in Biotic Interactions. In Secondary Metabolites in Soil Ecology Edited by Karlovsky P. Berlin, Heidelberg: Springer Berlin Heidelberg; 2008: 107–126.[Soil Biology].
-
- Hamed MB, Karamanou S, Olafsdottir S, Basilio JSM, Simoens K, Tsolis KC, Van Mellaert L, Guethmundsdottir EE, Hreggvidsson GO, Anne J, et al. Large-scale production of a thermostable Rhodothermus marinus cellulase by heterologous secretion from Streptomyces lividans. Microb Cell Fact. 2017;16:232. doi: 10.1186/s12934-017-0847-x. - DOI - PMC - PubMed
-
- Lammertyn E, Van Mellaert L, Schacht S, Dillen C, Sablon E, Van Broekhoven A, Anne J. Evaluation of a novel subtilisin inhibitor gene and mutant derivatives for the expression and secretion of mouse Tumor necrosis factor alpha by Streptomyces lividans. Appl Environ Microbiol. 1997;63:1808–13. doi: 10.1128/aem.63.5.1808-1813.1997. - DOI - PMC - PubMed
-
- Pozidis C, Lammertyn E, Politou AS, Anne J, Tsiftsoglou AS, Sianidis G, Economou A. Protein secretion biotechnology using Streptomyces lividans: large-scale production of functional trimeric Tumor necrosis factor alpha. Biotechnol Bioeng. 2001;72:611–9. doi: 10.1002/1097-0290(20010320)72:6<611::AID-BIT1026>3.0.CO;2-0. - DOI - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
