

An acyl-CoA thioesterase is essential for the biosynthesis of a key dauer pheromone in C. elegans
- PMID: 38183989
- PMCID: PMC11102344
- DOI: 10.1016/j.chembiol.2023.12.006
An acyl-CoA thioesterase is essential for the biosynthesis of a key dauer pheromone in C. elegans
Abstract
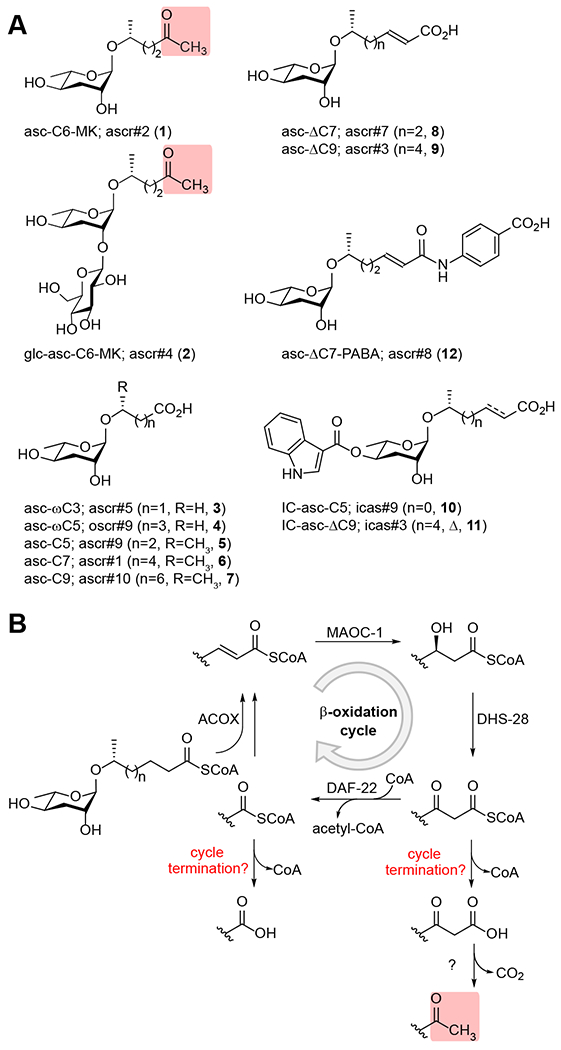
Methyl ketone (MK)-ascarosides represent essential components of several pheromones in Caenorhabditis elegans, including the dauer pheromone, which triggers the stress-resistant dauer larval stage, and the male-attracting sex pheromone. Here, we identify an acyl-CoA thioesterase, ACOT-15, that is required for the biosynthesis of MK-ascarosides. We propose a model in which ACOT-15 hydrolyzes the β-keto acyl-CoA side chain of an ascaroside intermediate during β-oxidation, leading to decarboxylation and formation of the MK. Using comparative metabolomics, we identify additional ACOT-15-dependent metabolites, including an unusual piperidyl-modified ascaroside, reminiscent of the alkaloid pelletierine. The β-keto acid generated by ACOT-15 likely couples to 1-piperideine to produce the piperidyl ascaroside, which is much less dauer-inducing than the dauer pheromone, asc-C6-MK (ascr#2, 1). The bacterial food provided influences production of the piperidyl ascaroside by the worm. Our work shows how the biosynthesis of MK- and piperidyl ascarosides intersect and how bacterial food may impact chemical signaling in the worm.
Keywords: ACOT-15; Caenorhabditis elegans; acyl-CoA thioesterase; anaferine; ascaroside; ascarosides; ascr#2; attraction; avoidance; beta-oxidation; dauer; hydroxylamine; lifespan; metabolomics; methyl ketone; natural products; pelletierine; pheromone; piperideine; piperidine; structure elucidation.
Copyright © 2023 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





Similar articles
-
Biosynthetic tailoring of existing ascaroside pheromones alters their biological function in C. elegans.Elife. 2018 Jun 4;7:e33286. doi: 10.7554/eLife.33286. Elife. 2018. PMID: 29863473 Free PMC article.
-
Ascaroside expression in Caenorhabditis elegans is strongly dependent on diet and developmental stage.PLoS One. 2011 Mar 15;6(3):e17804. doi: 10.1371/journal.pone.0017804. PLoS One. 2011. PMID: 21423575 Free PMC article.
-
Structural characterization of acyl-CoA oxidases reveals a direct link between pheromone biosynthesis and metabolic state in Caenorhabditis elegans.Proc Natl Acad Sci U S A. 2016 Sep 6;113(36):10055-60. doi: 10.1073/pnas.1608262113. Epub 2016 Aug 22. Proc Natl Acad Sci U S A. 2016. PMID: 27551084 Free PMC article.
-
Ascaroside Pheromones: Chemical Biology and Pleiotropic Neuronal Functions.Int J Mol Sci. 2019 Aug 9;20(16):3898. doi: 10.3390/ijms20163898. Int J Mol Sci. 2019. PMID: 31405082 Free PMC article. Review.
-
Ascaroside signaling in C. elegans.WormBook. 2013 Jan 18:1-22. doi: 10.1895/wormbook.1.155.1. WormBook. 2013. PMID: 23355522 Free PMC article. Review.
Cited by
-
Comparative Metabolomics Identifies the Roles of Acyl-CoA Oxidases in the Biosynthesis of Ascarosides and a Complex Family of Secreted N-Acylethanolamines.ACS Chem Biol. 2025 Jun 20;20(6):1298-1308. doi: 10.1021/acschembio.5c00126. Epub 2025 Jun 5. ACS Chem Biol. 2025. PMID: 40468644
-
Biosynthesis of modular signaling molecules requires functional diversification of carboxylesterases in Pristionchus pacificus.Commun Biol. 2025 Aug 13;8(1):1213. doi: 10.1038/s42003-025-08641-4. Commun Biol. 2025. PMID: 40804495 Free PMC article.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
