

Silica-associated proteins from hexactinellid sponges support an alternative evolutionary scenario for biomineralization in Porifera
- PMID: 38185711
- PMCID: PMC10772126
- DOI: 10.1038/s41467-023-44226-7
Silica-associated proteins from hexactinellid sponges support an alternative evolutionary scenario for biomineralization in Porifera
Abstract
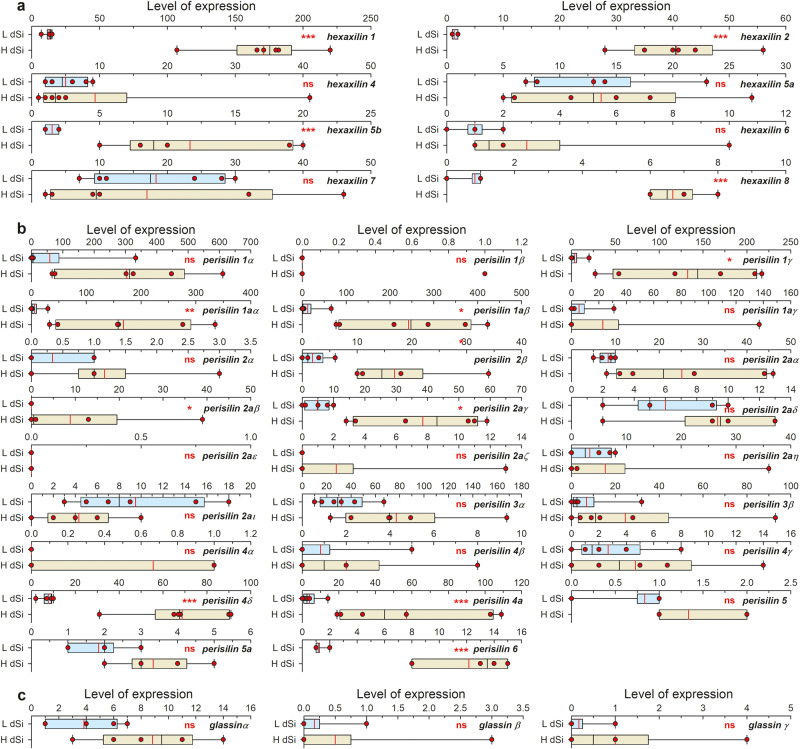
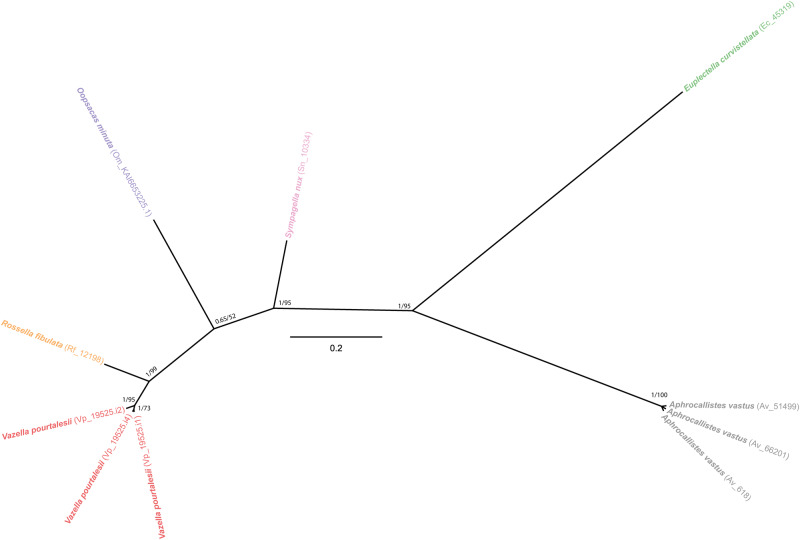
Metazoans use silicon traces but rarely develop extensive silica skeletons, except for the early-diverging lineage of sponges. The mechanisms underlying metazoan silicification remain incompletely understood, despite significant biotechnological and evolutionary implications. Here, the characterization of two proteins identified from hexactinellid sponge silica, hexaxilin and perisilin, supports that the three classes of siliceous sponges (Hexactinellida, Demospongiae, and Homoscleromorpha) use independent protein machineries to build their skeletons, which become non-homologous structures. Hexaxilin forms the axial filament to intracellularly pattern the main symmetry of the skeletal parts, while perisilin appears to operate in their thickening, guiding extracellular deposition of peripheral silica, as does glassin, a previously characterized hexactinellid silicifying protein. Distant hexaxilin homologs occur in some bilaterians with siliceous parts, suggesting putative conserved silicifying activity along metazoan evolution. The findings also support that ancestral Porifera were non-skeletonized, acquiring silica skeletons only after diverging into major classes, what reconciles molecular-clock dating and the fossil record.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







Similar articles
-
Glassin, a histidine-rich protein from the siliceous skeletal system of the marine sponge Euplectella, directs silica polycondensation.Proc Natl Acad Sci U S A. 2015 Sep 15;112(37):11449-54. doi: 10.1073/pnas.1506968112. Epub 2015 Aug 10. Proc Natl Acad Sci U S A. 2015. PMID: 26261346 Free PMC article.
-
The analysis of eight transcriptomes from all poriferan classes reveals surprising genetic complexity in sponges.Mol Biol Evol. 2014 May;31(5):1102-20. doi: 10.1093/molbev/msu057. Epub 2014 Feb 4. Mol Biol Evol. 2014. PMID: 24497032
-
The genome of the reef-building glass sponge Aphrocallistes vastus provides insights into silica biomineralization.R Soc Open Sci. 2023 Jun 21;10(6):230423. doi: 10.1098/rsos.230423. eCollection 2023 Jun. R Soc Open Sci. 2023. PMID: 37351491 Free PMC article.
-
Deep phylogeny and evolution of sponges (phylum Porifera).Adv Mar Biol. 2012;61:1-78. doi: 10.1016/B978-0-12-387787-1.00007-6. Adv Mar Biol. 2012. PMID: 22560777 Review.
-
Siliceous spicules and skeleton frameworks in sponges: origin, diversity, ultrastructural patterns, and biological functions.Microsc Res Tech. 2003 Nov 1;62(4):279-99. doi: 10.1002/jemt.10395. Microsc Res Tech. 2003. PMID: 14534903 Review.
Cited by
-
Mineral Composition of Skeletal Elements in Dorid Nudibranchia Onchidoris muricata (Gastropoda, Mollusca).Biomimetics (Basel). 2025 Mar 29;10(4):211. doi: 10.3390/biomimetics10040211. Biomimetics (Basel). 2025. PMID: 40277610 Free PMC article.
-
A late-Ediacaran crown-group sponge animal.Nature. 2024 Jun;630(8018):905-911. doi: 10.1038/s41586-024-07520-y. Epub 2024 Jun 5. Nature. 2024. PMID: 38839967
-
Silactins and Structural Diversity of Biosilica in Sponges.Biomimetics (Basel). 2024 Jun 27;9(7):393. doi: 10.3390/biomimetics9070393. Biomimetics (Basel). 2024. PMID: 39056834 Free PMC article.
-
Spongy but not glassy: Ediacaran fossil provides insight into early animal evolution.Nature. 2024 Jun 26. doi: 10.1038/d41586-024-02033-0. Online ahead of print. Nature. 2024. PMID: 38926561 No abstract available.
-
Biominerals and Bioinspired Materials in Biosensing: Recent Advancements and Applications.Int J Mol Sci. 2024 Apr 25;25(9):4678. doi: 10.3390/ijms25094678. Int J Mol Sci. 2024. PMID: 38731897 Free PMC article. Review.
References
-
- Sarikaya M, et al. Biomimetic model of a sponge-spicular optical fiber—mechanical properties and structure. J. Mater. Res. 2001;16:1420–1428. doi: 10.1557/JMR.2001.0198. - DOI
-
- Ehrlich H, et al. Supercontinuum generation in naturally occurring glass sponges spicules. Adv. Opt. Mater. 2016;4:1608–1613. doi: 10.1002/adom.201600454. - DOI
-
- Zhang M, et al. Robust and scalable in vitro surface mineralization of inert polymers with a rationally designed molecular bridge. ACS Appl. Mater. Interfaces. 2022;22:25. - PubMed
-
- Walter SL, Flinn BD, Mayer G. Mechanisms of toughening of a natural rigid composite. Mater. Sci. Eng. C. 2007;27:570–574. doi: 10.1016/j.msec.2006.05.020. - DOI
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
