

This is a preprint.
PRDM3/16 Regulate Chromatin Accessibility Required for NKX2-1 Mediated Alveolar Epithelial Differentiation and Function
- PMID: 38187557
- PMCID: PMC10769259
- DOI: 10.1101/2023.12.20.570481
PRDM3/16 Regulate Chromatin Accessibility Required for NKX2-1 Mediated Alveolar Epithelial Differentiation and Function
Update in
-
PRDM3/16 regulate chromatin accessibility required for NKX2-1 mediated alveolar epithelial differentiation and function.Nat Commun. 2024 Sep 16;15(1):8112. doi: 10.1038/s41467-024-52154-3. Nat Commun. 2024. PMID: 39284798 Free PMC article.
Abstract
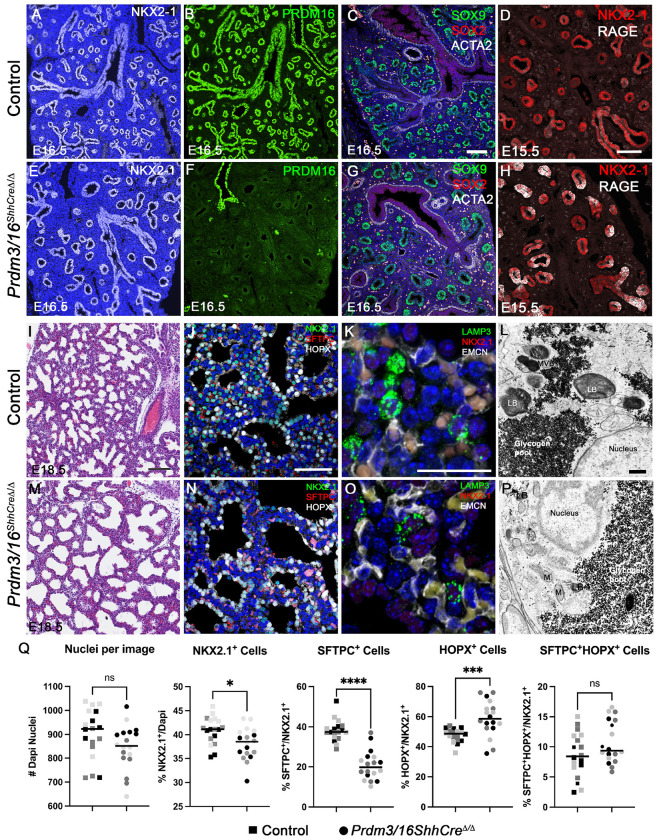
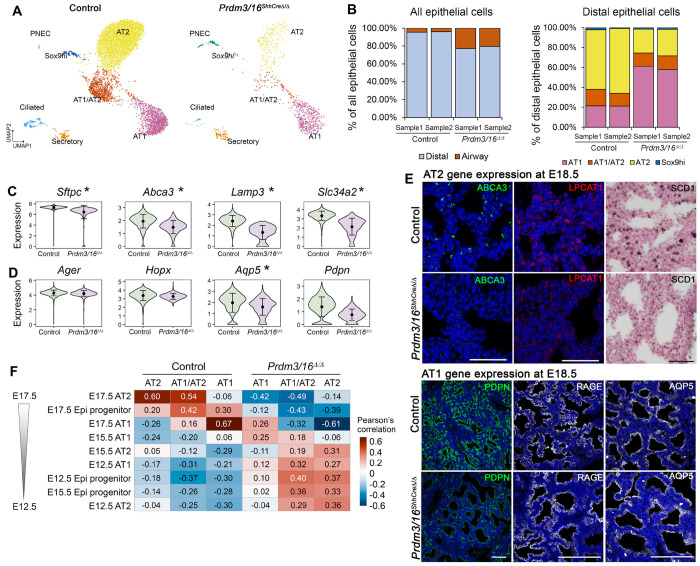
Differential chromatin accessibility accompanies and mediates transcriptional control of diverse cell fates and their differentiation during embryogenesis. While the critical role of NKX2-1 and its transcriptional targets in lung morphogenesis and pulmonary epithelial cell differentiation is increasingly known, mechanisms by which chromatin accessibility alters the epigenetic landscape and how NKX2-1 interacts with other co-activators required for alveolar epithelial cell differentiation and function are not well understood. Here, we demonstrate that the paired domain zinc finger transcriptional regulators PRDM3 and PRDM16 regulate chromatin accessibility to mediate cell differentiation decisions during lung morphogenesis. Combined deletion of Prdm3 and Prdm16 in early lung endoderm caused perinatal lethality due to respiratory failure from loss of AT2 cell function. Prdm3/16 deletion led to the accumulation of partially differentiated AT1 cells and loss of AT2 cells. Combination of single cell RNA-seq, bulk ATAC-seq, and CUT&RUN demonstrated that PRDM3 and PRDM16 enhanced chromatin accessibility at NKX2-1 transcriptional targets in peripheral epithelial cells, all three factors binding together at a multitude of cell-type specific cis-active DNA elements. Network analysis demonstrated that PRDM3/16 regulated genes critical for perinatal AT2 cell differentiation, surfactant homeostasis, and innate host defense. Lineage specific deletion of PRDM3/16 in AT2 cells led to lineage infidelity, with PRDM3/16 null cells acquiring partial AT1 fate. Together, these data demonstrate that NKX2-1-dependent regulation of alveolar epithelial cell differentiation is mediated by epigenomic modulation via PRDM3/16.
Keywords: Alveolar Epithelial Cell; Chromatin Accessibility; Differentiation; PRDM; Pulmonary Surfactant.
Conflict of interest statement
Competing interests: The Authors declare that they have no competing interests for the current work, including patents, financial holdings, advisory positions, or other interests.
Figures




References
-
- Kimura S., et al. , The T/ebp null mouse: thyroid-specific enhancer-binding protein is essential for the organogenesis of the thyroid, lung, ventral forebrain, and pituitary. Genes Dev, 1996. 10(1): p. 60–9. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials