

This is a preprint.
Interferon signaling in the nasal epithelium distinguishes among lethal and common cold respiratory viruses and is critical for viral clearance
- PMID: 38187597
- PMCID: PMC10769301
- DOI: 10.1101/2023.12.18.571720
Interferon signaling in the nasal epithelium distinguishes among lethal and common cold respiratory viruses and is critical for viral clearance
Update in
-
Interferon signaling in the nasal epithelium distinguishes among lethal and common cold coronaviruses and mediates viral clearance.Proc Natl Acad Sci U S A. 2024 May 21;121(21):e2402540121. doi: 10.1073/pnas.2402540121. Epub 2024 May 17. Proc Natl Acad Sci U S A. 2024. PMID: 38758698 Free PMC article.
Abstract
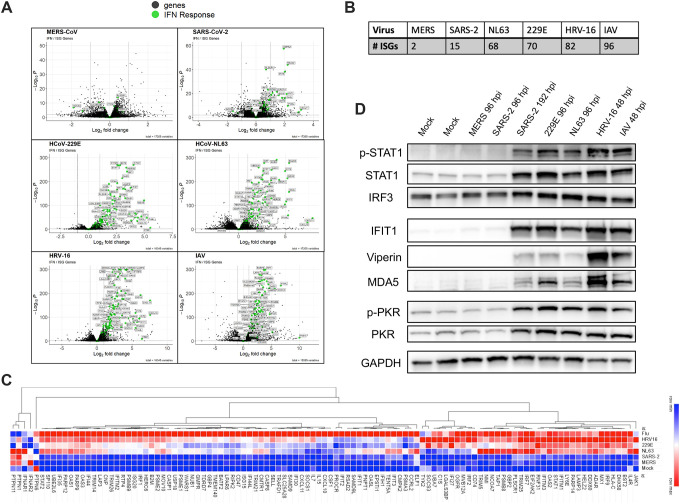
All respiratory viruses establish primary infections in the nasal epithelium, where efficient innate immune induction may prevent dissemination to the lower airway and thus minimize pathogenesis. Human coronaviruses (HCoVs) cause a range of pathologies, but the host and viral determinants of disease during common cold versus lethal HCoV infections are poorly understood. We model the initial site of infection using primary nasal epithelial cells cultured at air-liquid interface (ALI). HCoV-229E, HCoV-NL63 and human rhinovirus-16 are common cold-associated viruses that exhibit unique features in this model: early induction of antiviral interferon (IFN) signaling, IFN-mediated viral clearance, and preferential replication at nasal airway temperature (33°C) which confers muted host IFN responses. In contrast, lethal SARS-CoV-2 and MERS-CoV encode antagonist proteins that prevent IFN-mediated clearance in nasal cultures. Our study identifies features shared among common cold-associated viruses, highlighting nasal innate immune responses as predictive of infection outcomes and nasally-directed IFNs as potential therapeutics.
Keywords: common cold; coronavirus; interferon signaling; nasal epithelium; virus, temperature.
Conflict of interest statement
Competing Interest / Disclosures Statement: Susan R Weiss is on the Scientific Advisory Board of Ocugen, Inc. and consults for Powell Gilbert LLP. Noam A Cohen consults for GSK, AstraZeneca, Novartis, Sanofi/Regeneron; has US Patent “Therapy and Diagnostics for Respiratory Infection” (10,881,698 B2, WO20913112865) and a licensing agreement with GeneOne Life Sciences.
Figures






References
-
- Booth T.F., Kournikakis B., Bastien N., Ho J., Kobasa D., Stadnyk L., Li Y., Spence M., Paton S., Henry B., et al. (2005). Detection of airborne severe acute respiratory syndrome (SARS) coronavirus and environmental contamination in SARS outbreak units. Journal of Infectious Diseases 191, 1472–1477. 10.1086/429634. - DOI - PMC - PubMed
-
- Farzal Z., Basu S., Burke A., Fasanmade O.O., Lopez E.M., Bennett W.D., Ebert C.S., Zanation A.M., Senior B.A., and Kimbell J.S. (2019). Comparative study of simulated nebulized and spray particle deposition in chronic rhinosinusitis patients. Int Forum Allergy Rhinol 9, 746–758. 10.1002/alr.22324. - DOI - PMC - PubMed
-
- Dewitte-Orr S.J., and Mossman K.L. (2010). DsRNA and the innate antiviral immune response. Future Virol 5, 325–341. 10.2217/fvl.10.18. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous