

This is a preprint.
A Reinterpretation of the Relationship Between Persistent and Resurgent Sodium Currents
- PMID: 38187680
- PMCID: PMC10769191
- DOI: 10.1101/2023.10.25.564042
A Reinterpretation of the Relationship Between Persistent and Resurgent Sodium Currents
Update in
-
A Reinterpretation of the Relationship between Persistent and Resurgent Sodium Currents.J Neurosci. 2024 Jul 17;44(29):e2396232024. doi: 10.1523/JNEUROSCI.2396-23.2024. J Neurosci. 2024. PMID: 38858080 Free PMC article.
Abstract
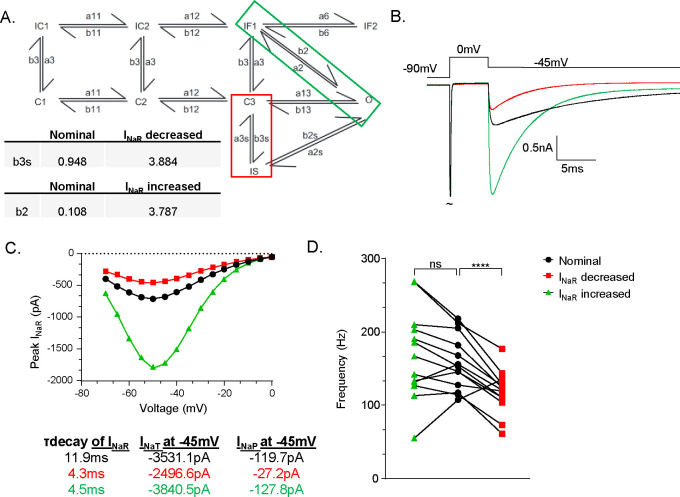
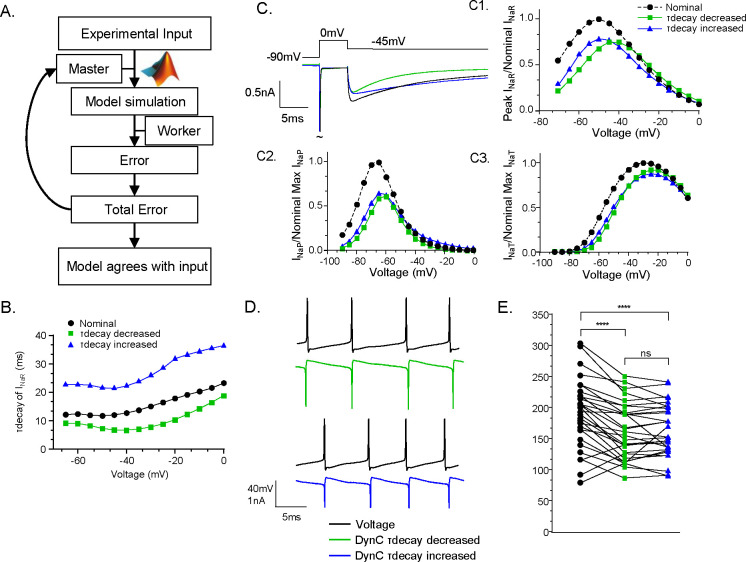
The resurgent sodium current (INaR) activates on membrane repolarization, such as during the downstroke of neuronal action potentials. Due to its unique activation properties, INaR is thought to drive high rates of repetitive neuronal firing. However, INaR is often studied in combination with the persistent or non-inactivating portion of sodium currents (INaP). We used dynamic clamp to test how INaR and INaP individually affect repetitive firing in adult cerebellar Purkinje neurons from male and female mice. We learned INaR does not scale repetitive firing rates due to its rapid decay at subthreshold voltages, and that subthreshold INaP is critical in regulating neuronal firing rate. Adjustments to the Nav conductance model used in these studies revealed INaP and INaR can be inversely scaled by adjusting occupancy in the slow inactivated kinetic state. Together with additional dynamic clamp experiments, these data suggest the regulation of sodium channel slow inactivation can fine-tune INaP and Purkinje neuron repetitive firing rates.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






References
-
- Aman T. K., & Raman I. M. (2010). Inwardly permeating Na ions generate the voltage dependence of resurgent Na current in cerebellar Purkinje neurons. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 30(16), 5629–5634. 10.1523/JNEUROSCI.0376-10.2010 - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials