

This is a preprint.
Neoantigen Cancer Vaccines and Different Immune Checkpoint Therapies Each Utilize Both Converging and Distinct Mechanisms that in Combination Enable Synergistic Therapeutic Efficacy
- PMID: 38187708
- PMCID: PMC10769249
- DOI: 10.1101/2023.12.20.570816
Neoantigen Cancer Vaccines and Different Immune Checkpoint Therapies Each Utilize Both Converging and Distinct Mechanisms that in Combination Enable Synergistic Therapeutic Efficacy
Update in
-
Comparing neoantigen cancer vaccines and immune checkpoint therapy unveils an effective vaccine and anti-TREM2 macrophage-targeting dual therapy.Cell Rep. 2024 Nov 26;43(11):114875. doi: 10.1016/j.celrep.2024.114875. Epub 2024 Oct 23. Cell Rep. 2024. PMID: 39446585 Free PMC article.
Abstract
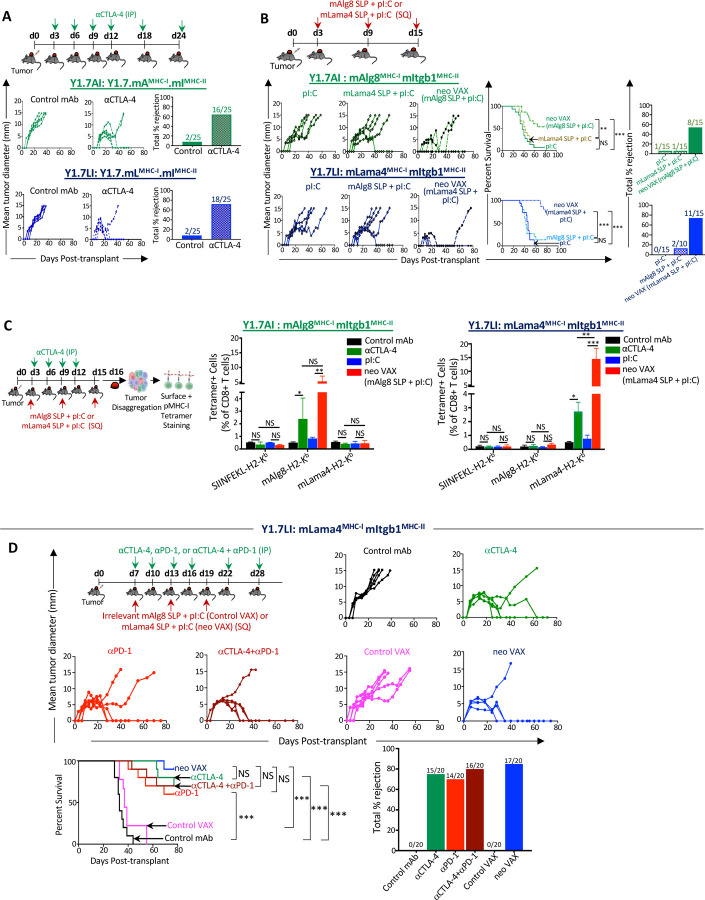
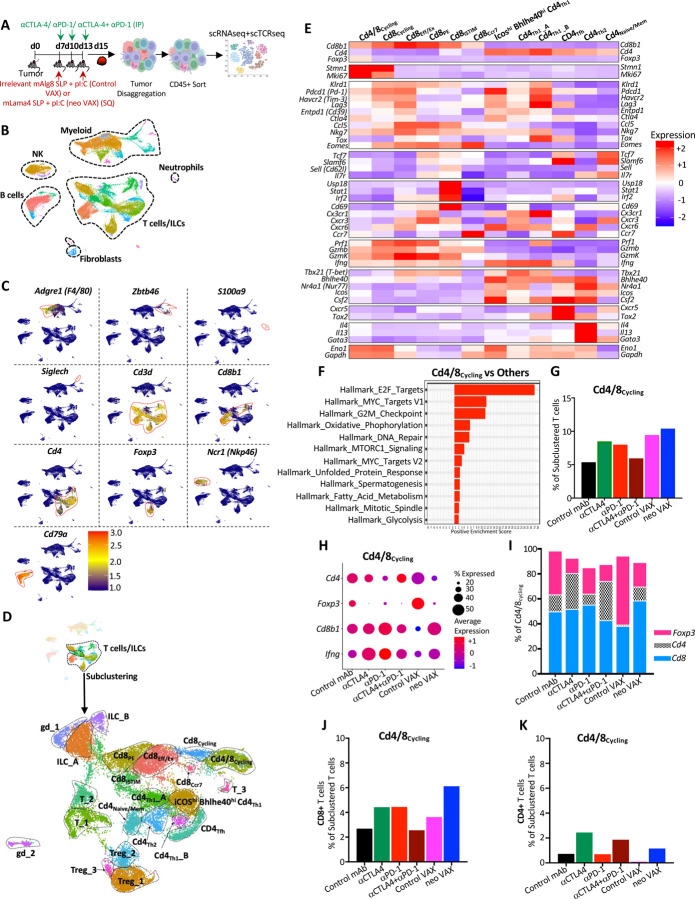
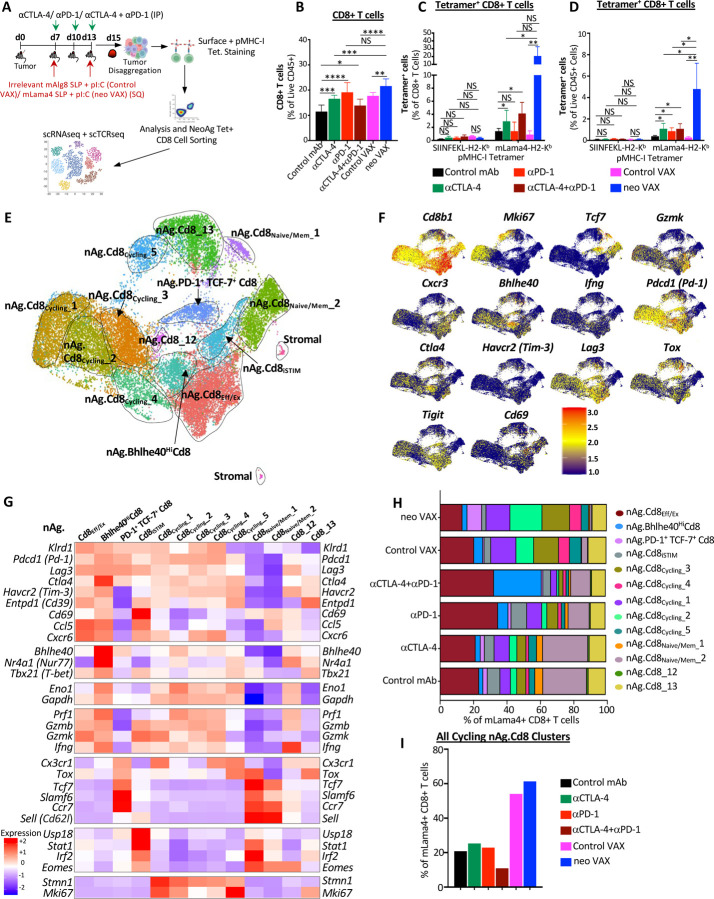
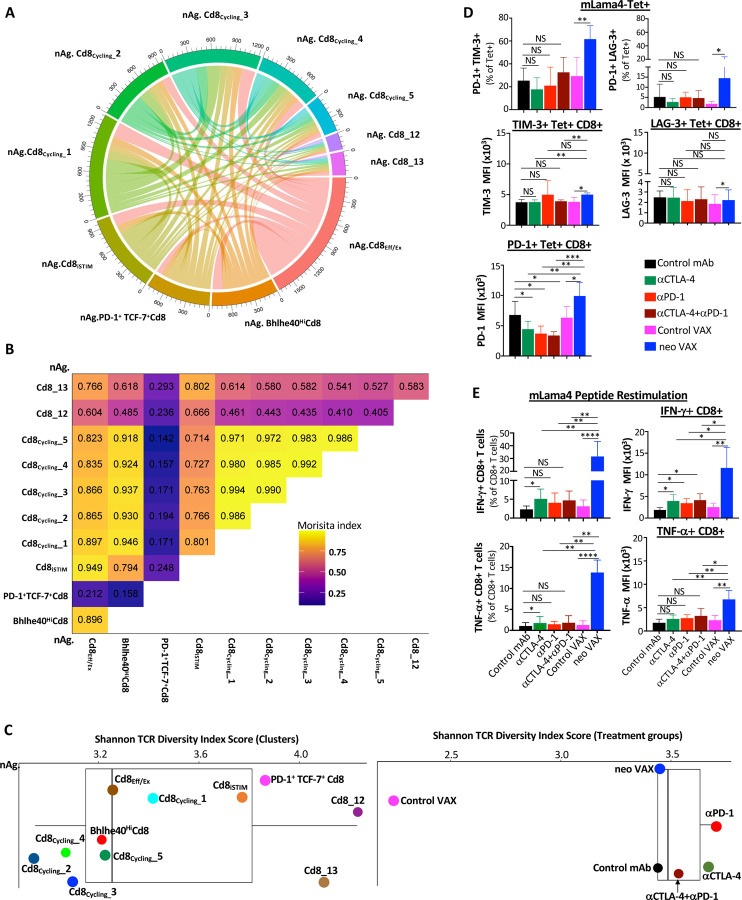
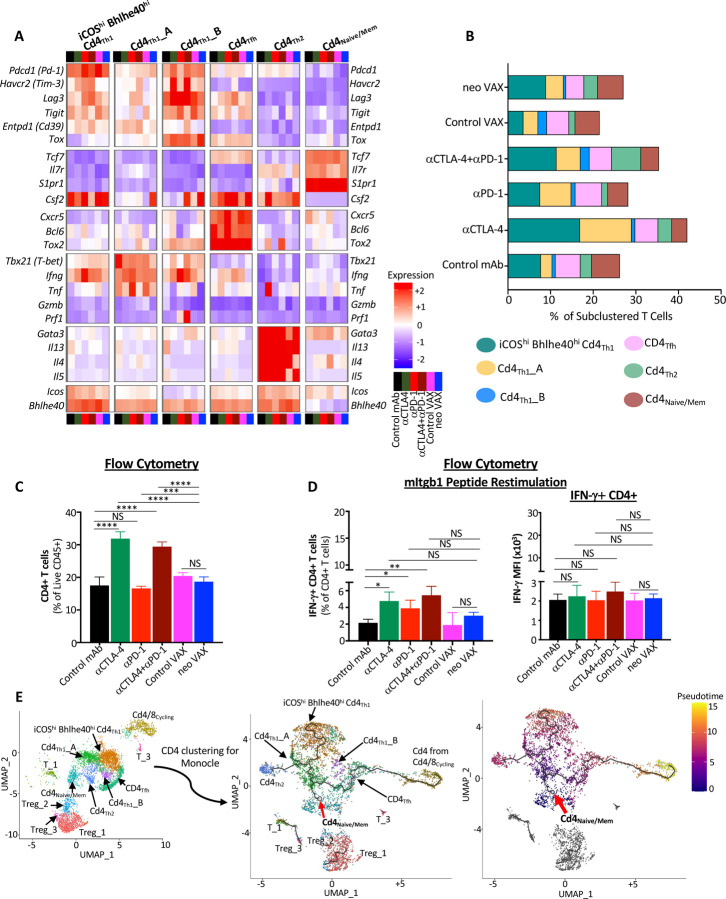
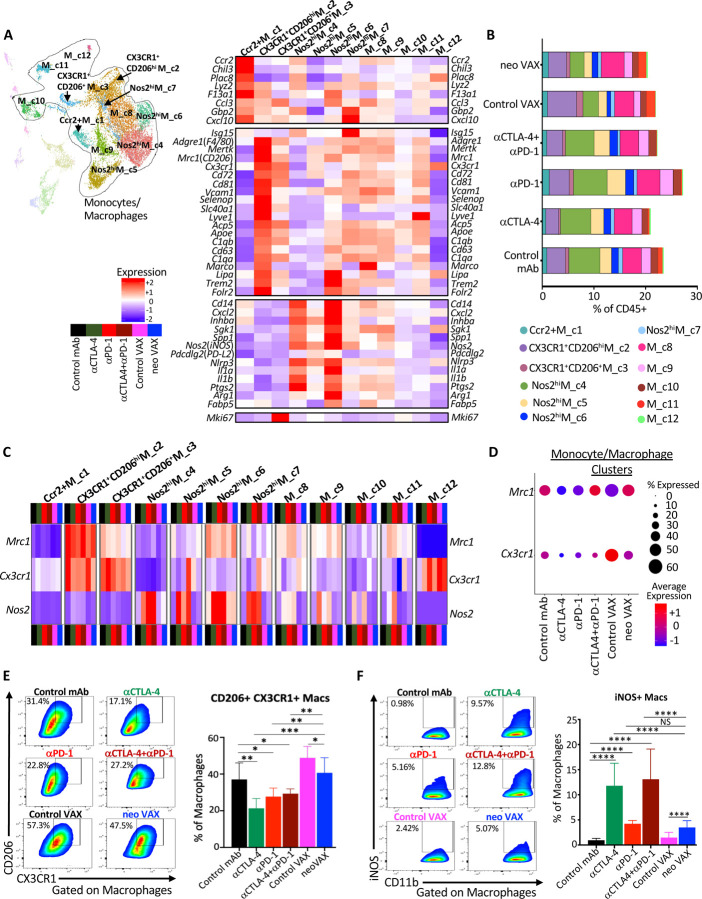
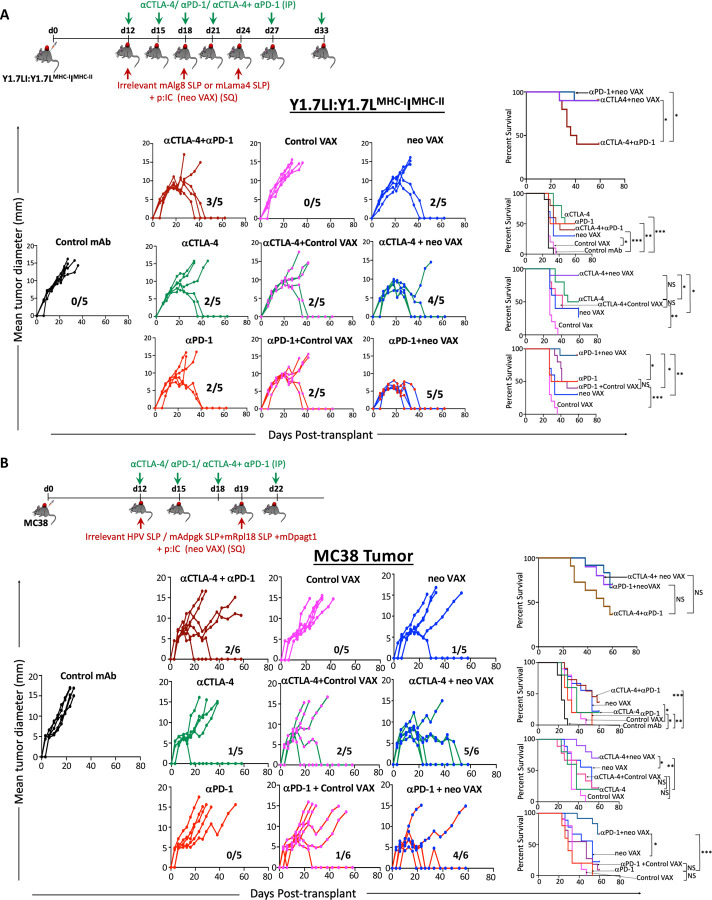
The goal of therapeutic cancer vaccines and immune checkpoint therapy (ICT) is to eliminate cancer by expanding and/or sustaining T cells with anti-tumor capabilities. However, whether cancer vaccines and ICT enhance anti-tumor immunity by distinct or overlapping mechanisms remains unclear. Here, we compared effective therapeutic tumor-specific mutant neoantigen (NeoAg) cancer vaccines with anti-CTLA-4 and/or anti-PD-1 ICT in preclinical models. Both NeoAg vaccines and ICT induce expansion of intratumoral NeoAg-specific CD8 T cells, though the degree of expansion and acquisition of effector activity was much more substantial following NeoAg vaccination. Further, we found that NeoAg vaccines are particularly adept at inducing proliferating and stem-like NeoAg-specific CD8 T cells. Single cell T cell receptor (TCR) sequencing revealed that TCR clonotype expansion and diversity of NeoAg-specific CD8 T cells relates to their phenotype and functional state associated with specific immunotherapies employed. Effective NeoAg vaccines and ICT required both CD8 and CD4 T cells. While NeoAg vaccines and anti-PD-1 affected the CD4 T cell compartment, it was to less of an extent than observed with anti-CTLA-4, which notably induced ICOS+Bhlhe40+ Th1-like CD4 T cells and, when combined with anti-PD-1, a small subset of Th2-like CD4 T cells. Although effective NeoAg vaccines or ICT expanded intratumoral M1-like iNOS+ macrophages, NeoAg vaccines expanded rather than suppressed (as observed with ICT) M2-like CX3CR1+CD206+ macrophages, associated with the vaccine adjuvant. Further, combining NeoAg vaccination with ICT induced superior efficacy compared to either therapy in isolation, highlighting the utility of combining these modalities to eliminate cancer.
Conflict of interest statement
Declaration of interests M.M. Gubin reports a personal honorarium of $1000.00 USD per year from Springer Nature Ltd for his role as an Associate Editor for the journal Nature Precision Oncology. No disclosures were reported by the other authors.
Figures
References
-
- Carreno B.M., Magrini V., Becker-Hapak M., Kaabinejadian S., Hundal J., Petti A.A., Ly A., Lie W.R., Hildebrand W.H., Mardis E.R., and Linette G.P. (2015). A dendritic cell vaccine increases the breadth and diversity of melanoma neoantigen-specific T cells. Science. 10.1126/science.aaa3828. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials