

This is a preprint.
CDCA7 is a hemimethylated DNA adaptor for the nucleosome remodeler HELLS
- PMID: 38187757
- PMCID: PMC10769307
- DOI: 10.1101/2023.12.19.572350
CDCA7 is a hemimethylated DNA adaptor for the nucleosome remodeler HELLS
Update in
-
CDCA7 is an evolutionarily conserved hemimethylated DNA sensor in eukaryotes.Sci Adv. 2024 Aug 23;10(34):eadp5753. doi: 10.1126/sciadv.adp5753. Epub 2024 Aug 23. Sci Adv. 2024. PMID: 39178260 Free PMC article.
Abstract
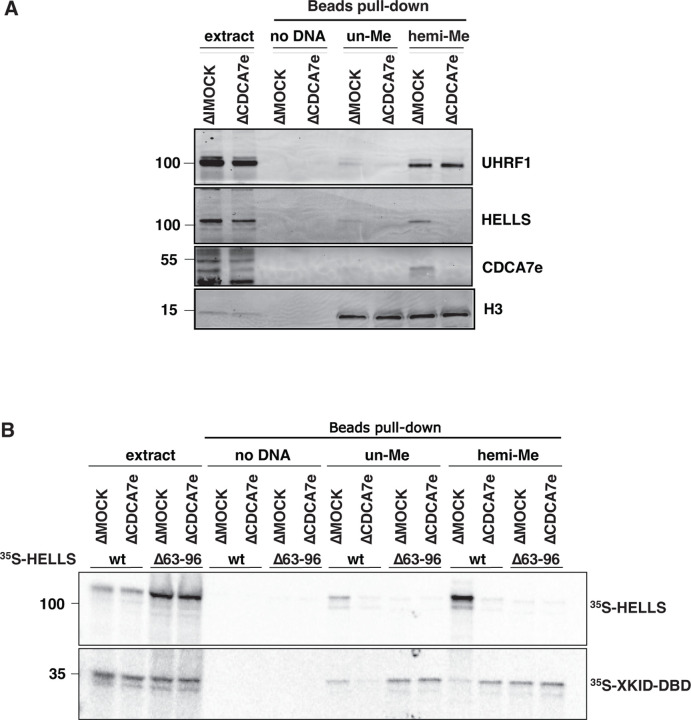
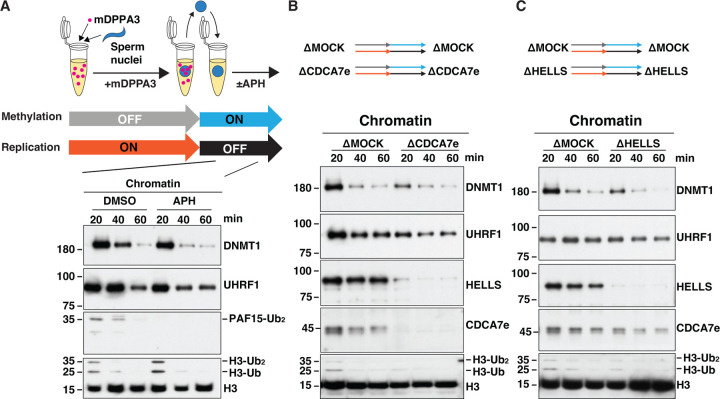
Mutations of the SNF2 family ATPase HELLS and its activator CDCA7 cause immunodeficiency-centromeric instability-facial anomalies (ICF) syndrome, characterized by hypomethylation at heterochromatin. The unique zinc-finger domain, zf-4CXXC_R1, of CDCA7 is widely conserved across eukaryotes but is absent from species that lack HELLS and DNA methyltransferases, implying its specialized relation with methylated DNA. Here we demonstrate that zf-4CXXC_R1 acts as a hemimethylated DNA sensor. The zf-4CXXC_R1 domain of CDCA7 selectively binds to DNA with a hemimethylated CpG, but not unmethylated or fully methylated CpG, and ICF disease mutations eliminated this binding. CDCA7 and HELLS interact via their N-terminal alpha helices, through which HELLS is recruited to hemimethylated DNA. While placement of a hemimethylated CpG within the nucleosome core particle can hinder its recognition by CDCA7, cryo-EM structure analysis of the CDCA7-nucleosome complex suggests that zf-4CXXC_R1 recognizes a hemimethylated CpG in the major groove at linker DNA. Our study provides insights into how the CDCA7-HELLS nucleosome remodeling complex uniquely assists maintenance DNA methylation.
Conflict of interest statement
Competing interests: H.F. is affiliated with Graduate School of Medical Sciences, Weill Cornell Medicine, and the Cell Biology Program at the Sloan Kettering Institute. The authors declare no competing interests.
Figures
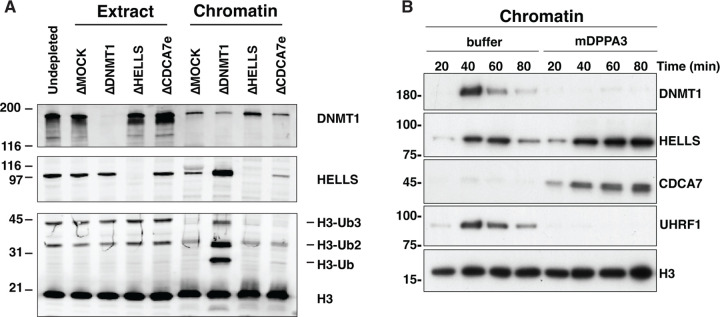
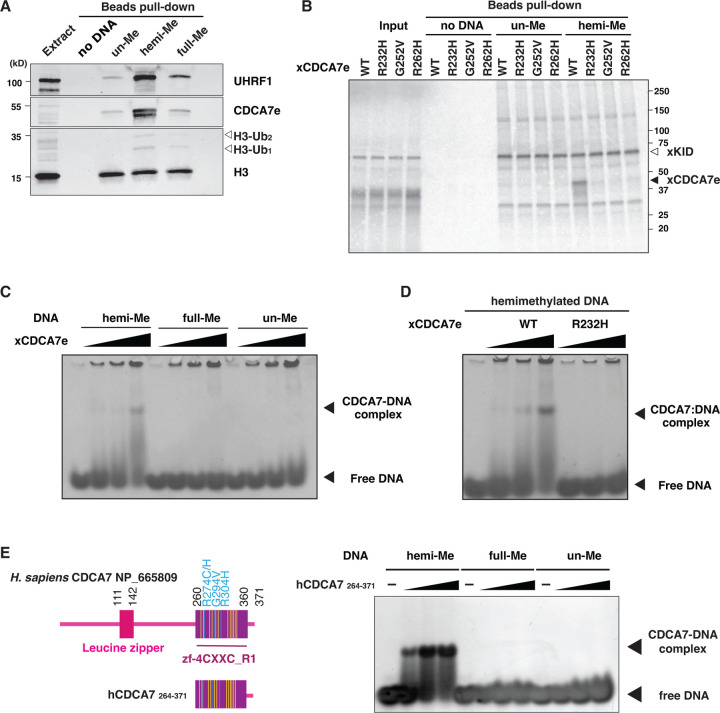
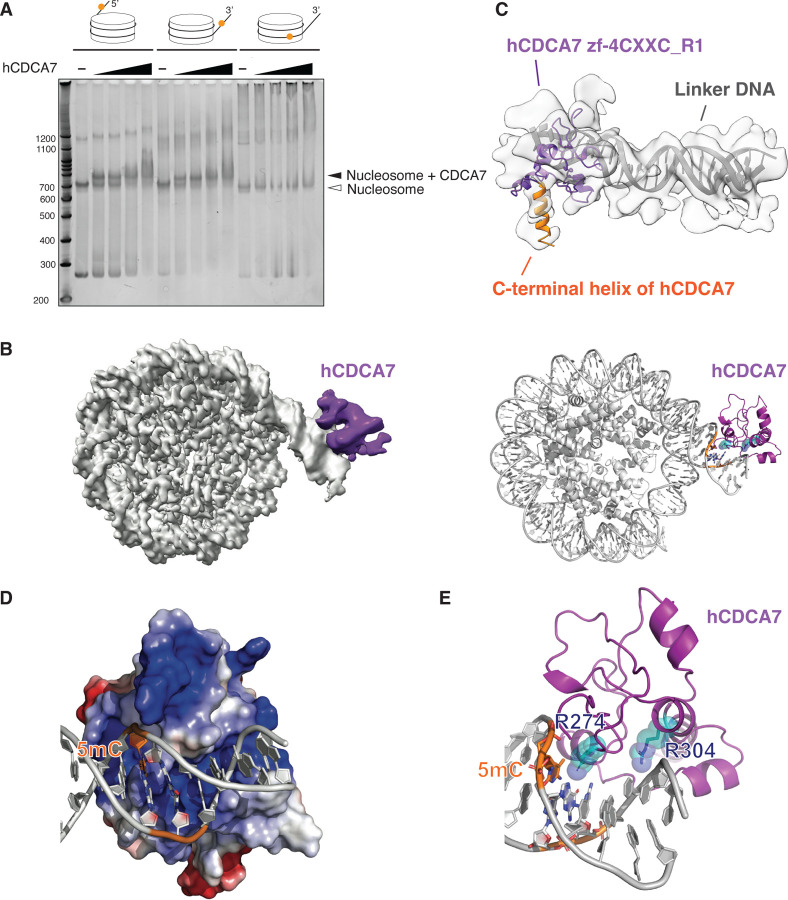
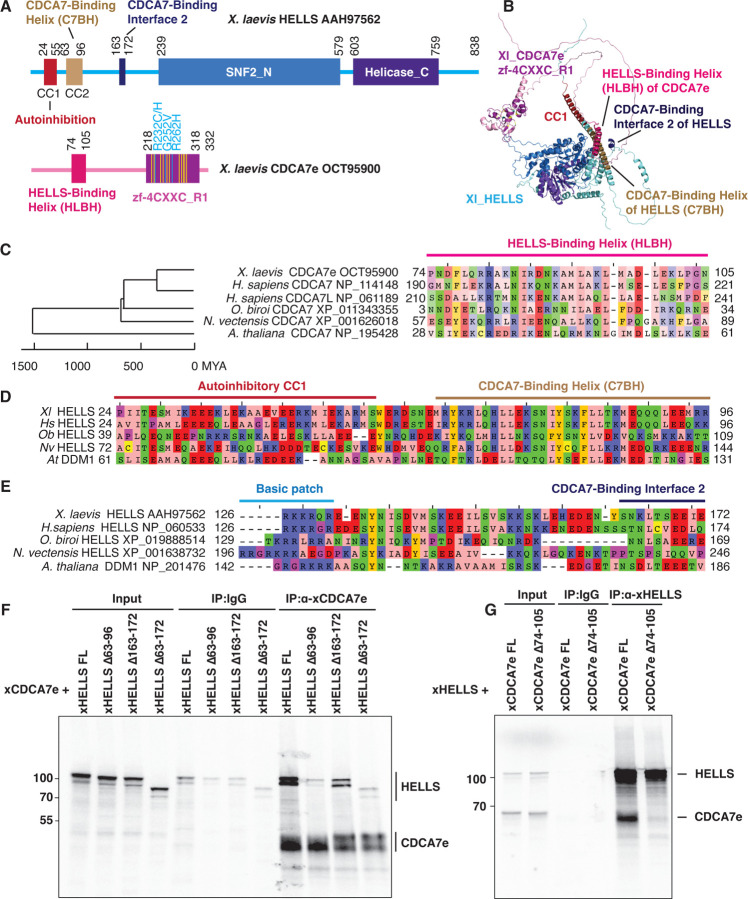
References
-
- Deniz O., Frost J. M., Branco M. R., Regulation of transposable elements by DNA modifications. Nat Rev Genet 20, 417–431 (2019). - PubMed
-
- Mattei A. L., Bailly N., Meissner A., DNA methylation: a historical perspective. Trends Genet 38, 676–707 (2022). - PubMed
-
- Miska E. A., Ferguson-Smith A. C., Transgenerational inheritance: Models and mechanisms of non-DNA sequence-based inheritance. Science 354, 59–63 (2016). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources