

Eomes-dependent mitochondrial regulation promotes survival of pathogenic CD4+ T cells during inflammation
- PMID: 38189779
- PMCID: PMC10772920
- DOI: 10.1084/jem.20230449
Eomes-dependent mitochondrial regulation promotes survival of pathogenic CD4+ T cells during inflammation
Abstract
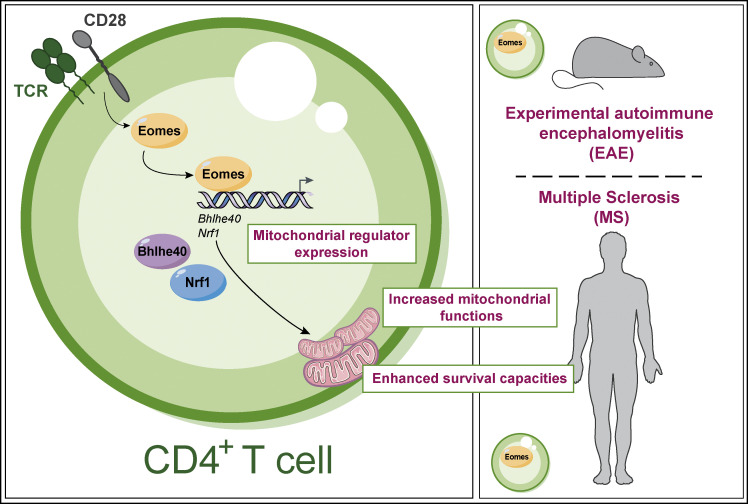
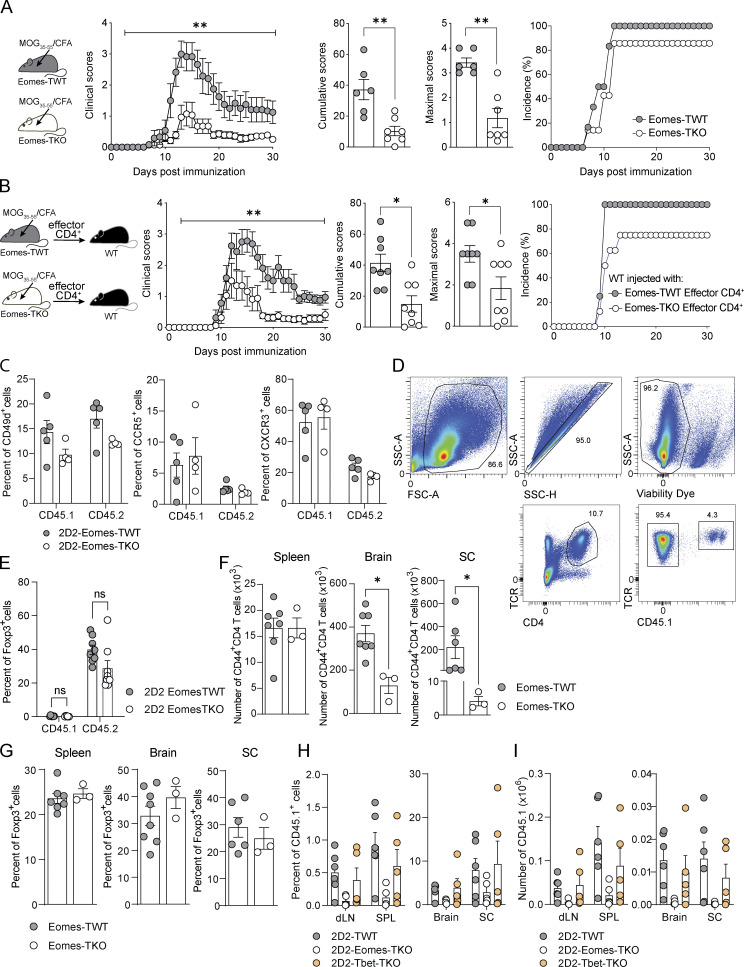
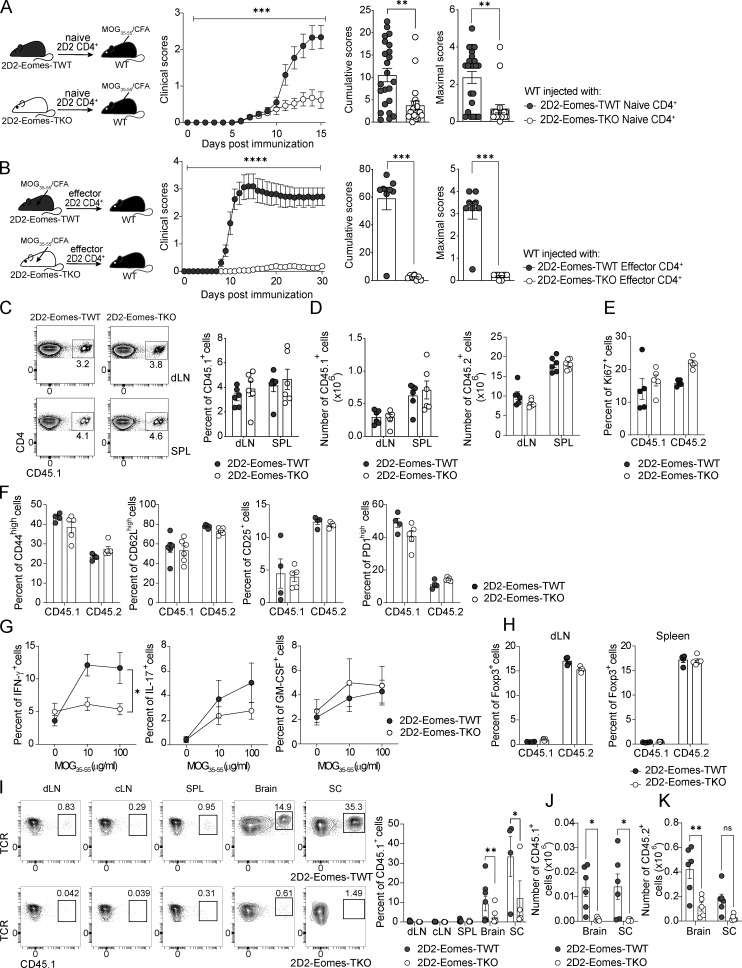
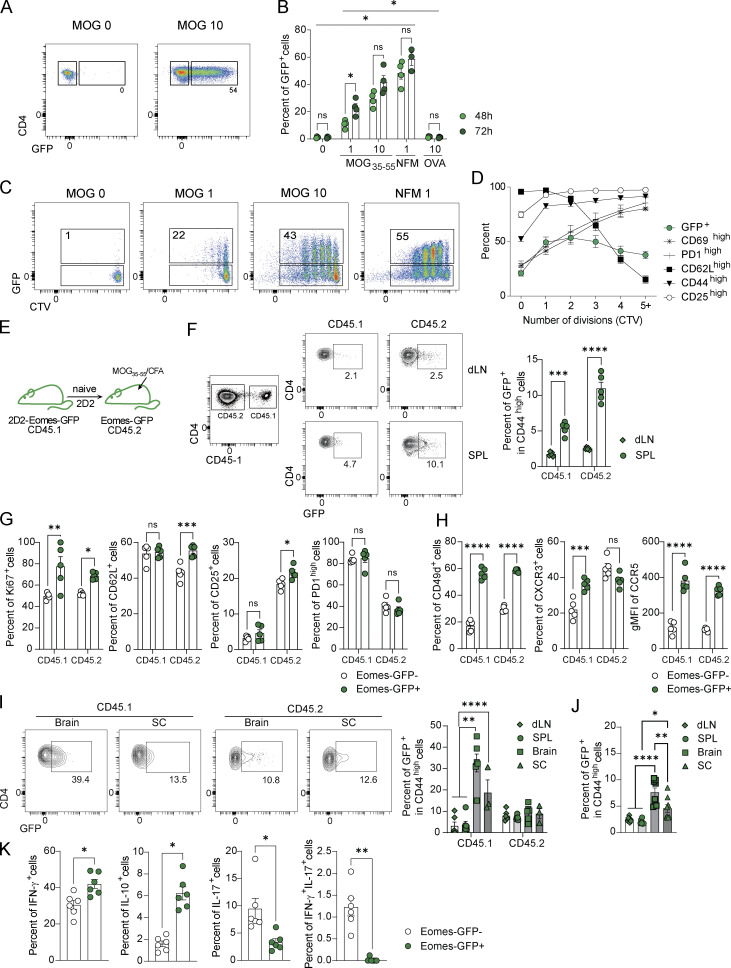
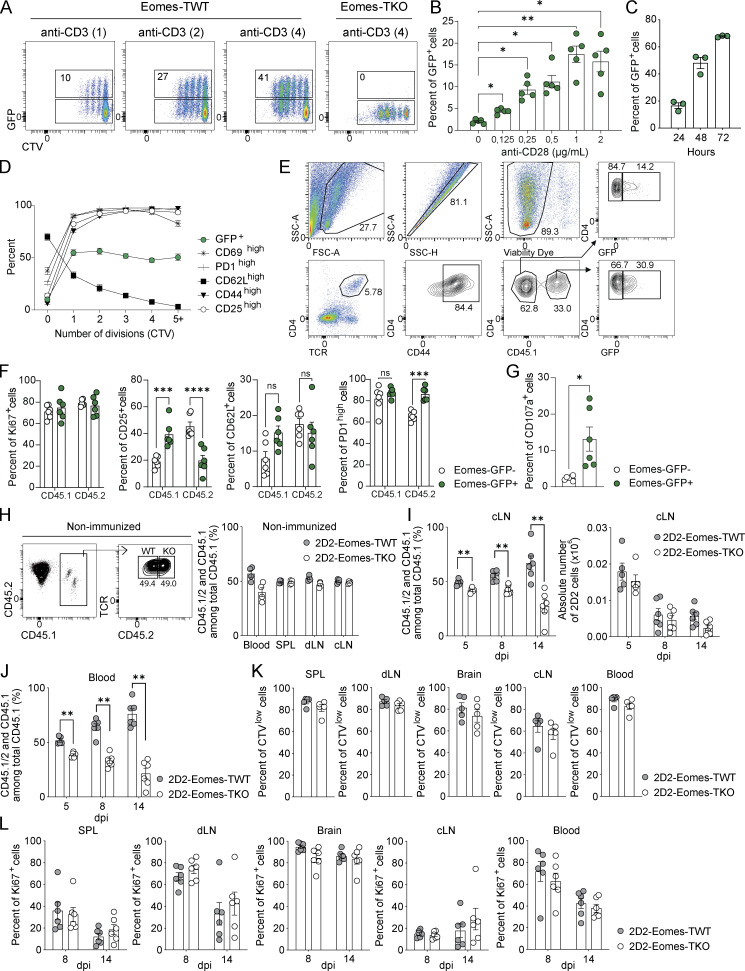
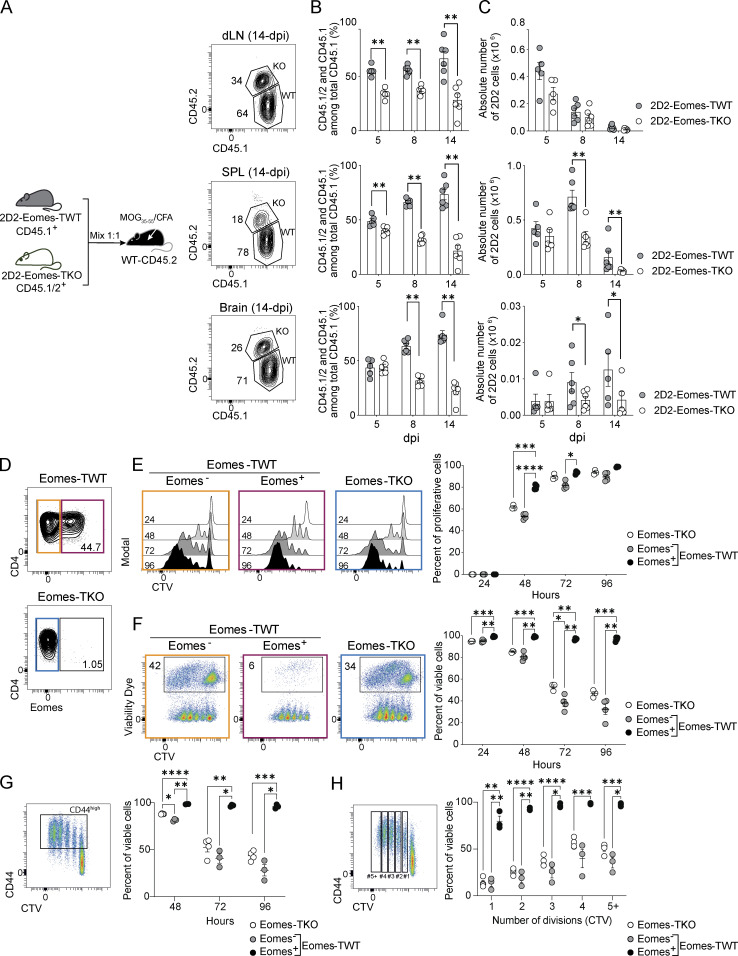
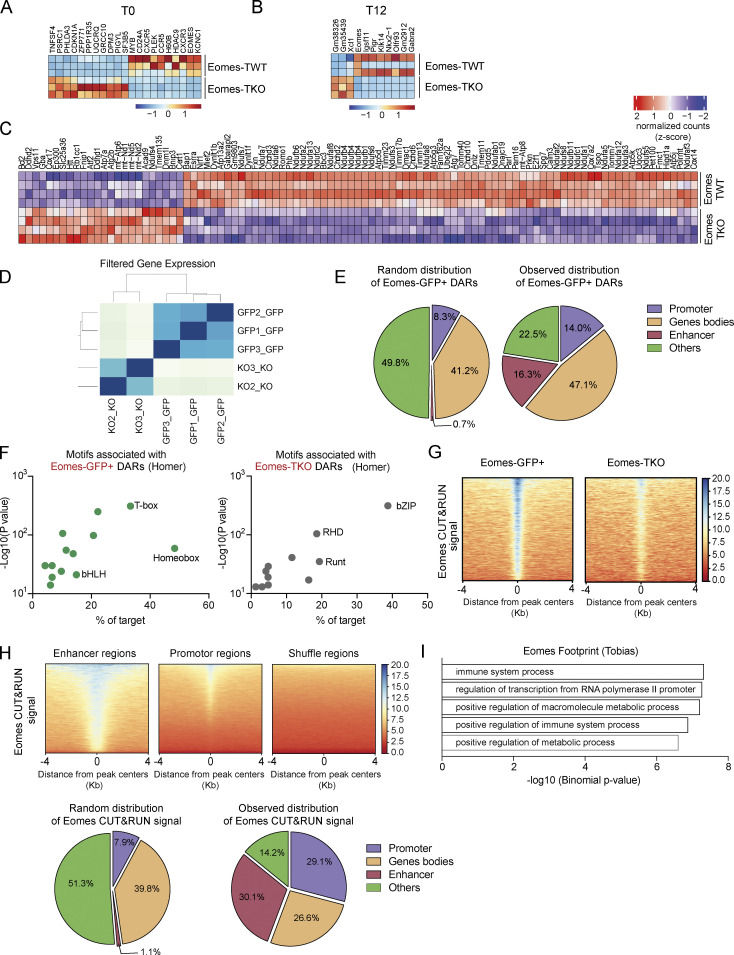
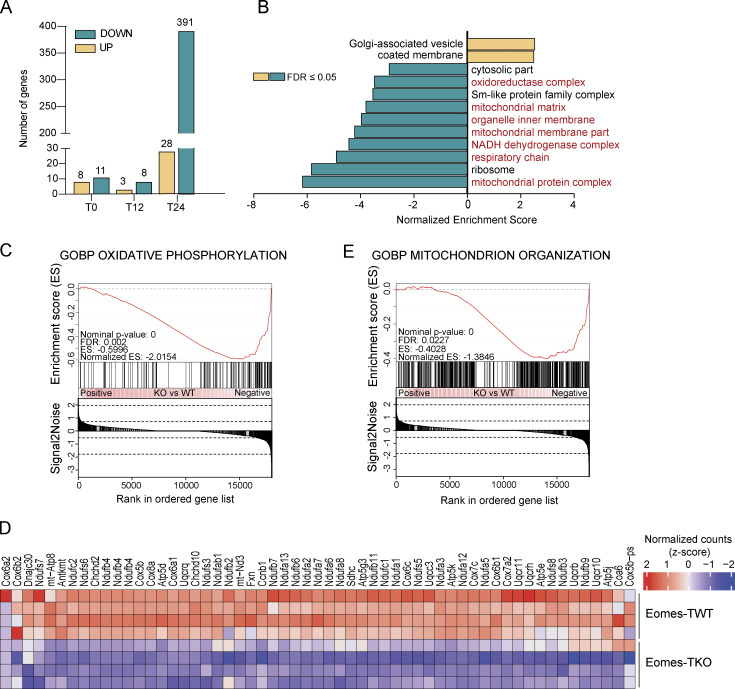
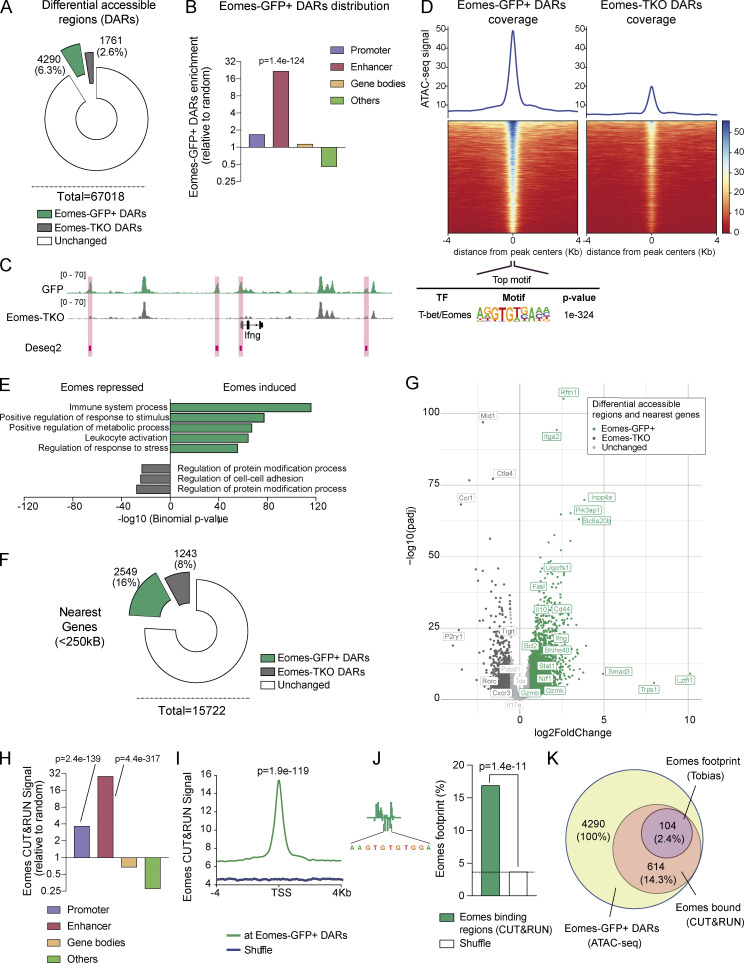
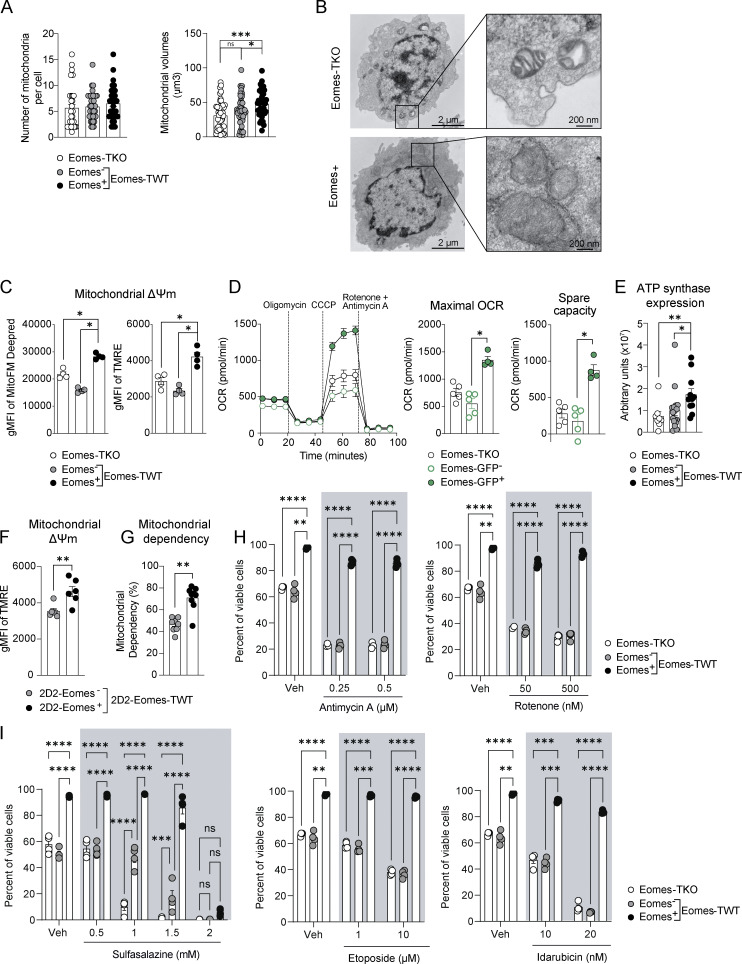
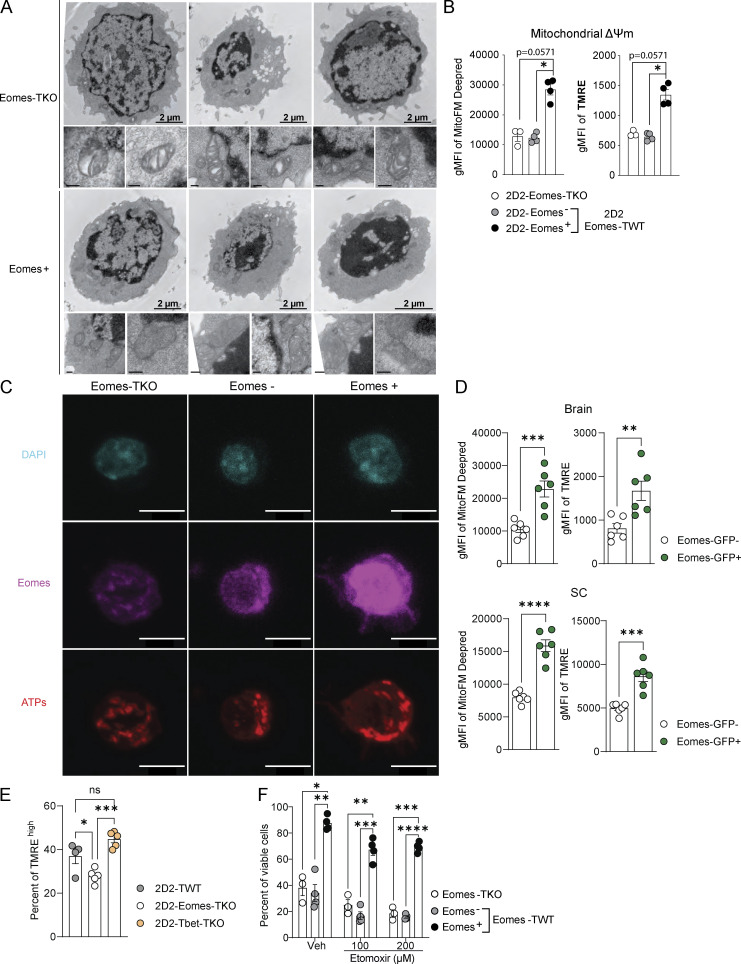
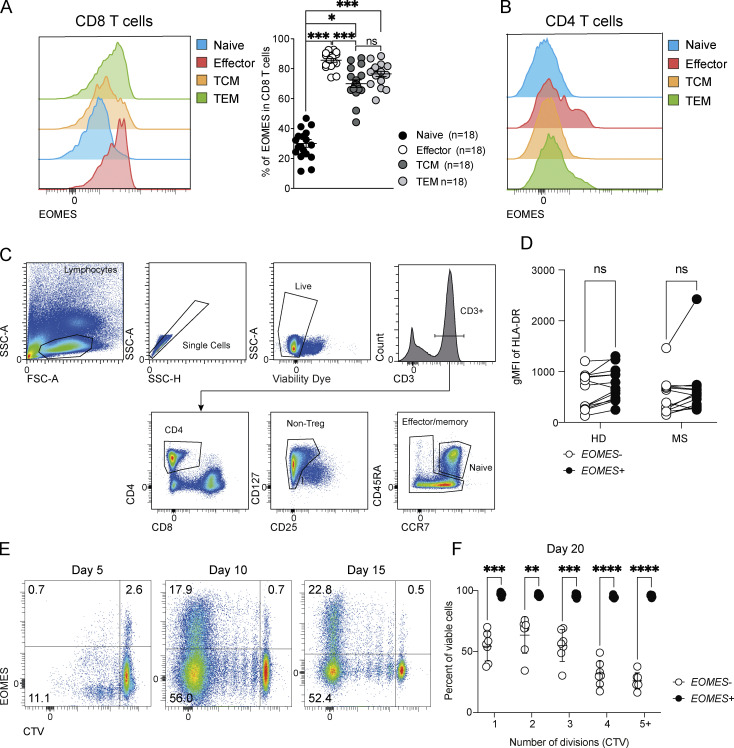
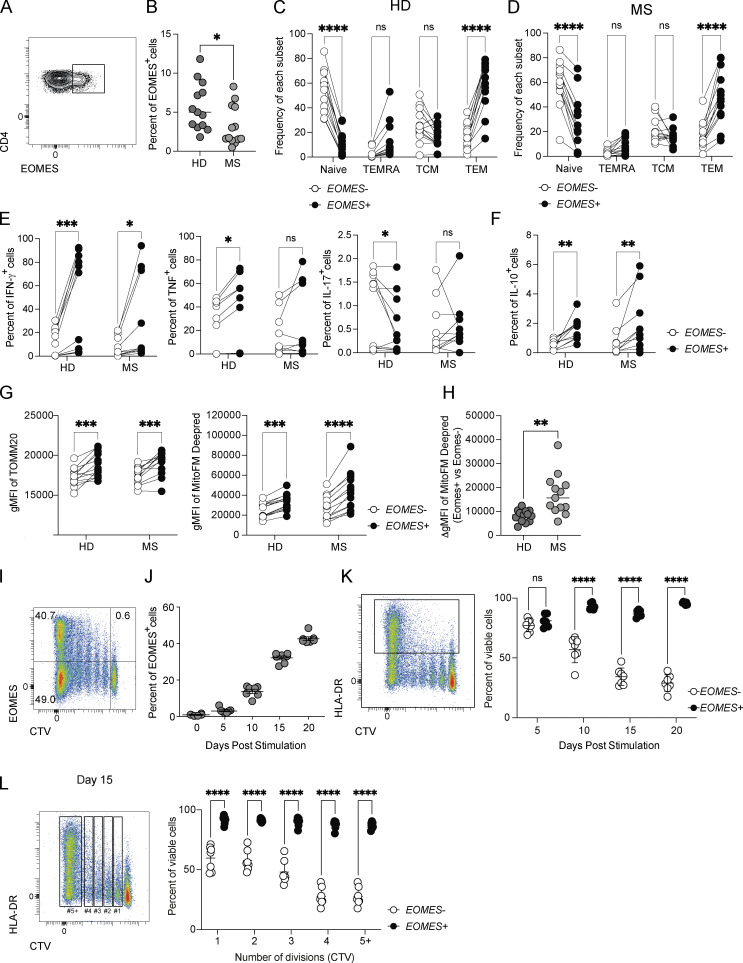
The mechanisms whereby Eomes controls tissue accumulation of T cells and strengthens inflammation remain ill-defined. Here, we show that Eomes deletion in antigen-specific CD4+ T cells is sufficient to protect against central nervous system (CNS) inflammation. While Eomes is dispensable for the initial priming of CD4+ T cells, it is required for long-term maintenance of CNS-infiltrating CD4+ T cells. We reveal that the impact of Eomes on effector CD4+ T cell longevity is associated with sustained expression of multiple genes involved in mitochondrial organization and functions. Accordingly, epigenetic studies demonstrate that Eomes supports mitochondrial function by direct binding to either metabolism-associated genes or mitochondrial transcriptional modulators. Besides, the significance of these findings was confirmed in CD4+ T cells from healthy donors and multiple sclerosis patients. Together, our data reveal a new mechanism by which Eomes promotes severity and chronicity of inflammation via the enhancement of CD4+ T cell mitochondrial functions and resistance to stress-induced cell death.
© 2024 Joulia et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures
References
-
- Adoue, V., Binet B., Malbec A., Fourquet J., Romagnoli P., van Meerwijk J.P.M., Amigorena S., and Joffre O.P.. 2019. The histone methyltransferase SETDB1 controls T Helper cell lineage integrity by repressing endogenous retroviruses. Immunity. 50:629–644.e8. 10.1016/j.immuni.2019.01.003 - DOI - PubMed
-
- Argüello, R.J., Combes A.J., Char R., Gigan J.P., Baaziz A.I., Bousiquot E., Camosseto V., Samad B., Tsui J., Yan P., et al. . 2020. SCENITH: A flow cytometry-based method to functionally profile energy metabolism with single-cell resolution. Cell Metab. 32:1063–1075.e7. 10.1016/j.cmet.2020.11.007 - DOI - PMC - PubMed
-
- Banerjee, A., Gordon S.M., Intlekofer A.M., Paley M.A., Mooney E.C., Lindsten T., Wherry E.J., and Reiner S.L.. 2010. Cutting edge: The transcription factor eomesodermin enables CD8+ T cells to compete for the memory cell niche. J. Immunol. 185:4988–4992. 10.4049/jimmunol.1002042 - DOI - PMC - PubMed
-
- Bentsen, M., Goymann P., Schultheis H., Klee K., Petrova A., Wiegandt R., Fust A., Preussner J., Kuenne C., Braun T., et al. . 2020. ATAC-seq footprinting unravels kinetics of transcription factor binding during zygotic genome activation. Nat. Commun. 11:4267. 10.1038/s41467-020-18035-1 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
