

Multiple pathways for glucose phosphate transport and utilization support growth of Cryptosporidium parvum
- PMID: 38191884
- PMCID: PMC10774378
- DOI: 10.1038/s41467-024-44696-3
Multiple pathways for glucose phosphate transport and utilization support growth of Cryptosporidium parvum
Abstract
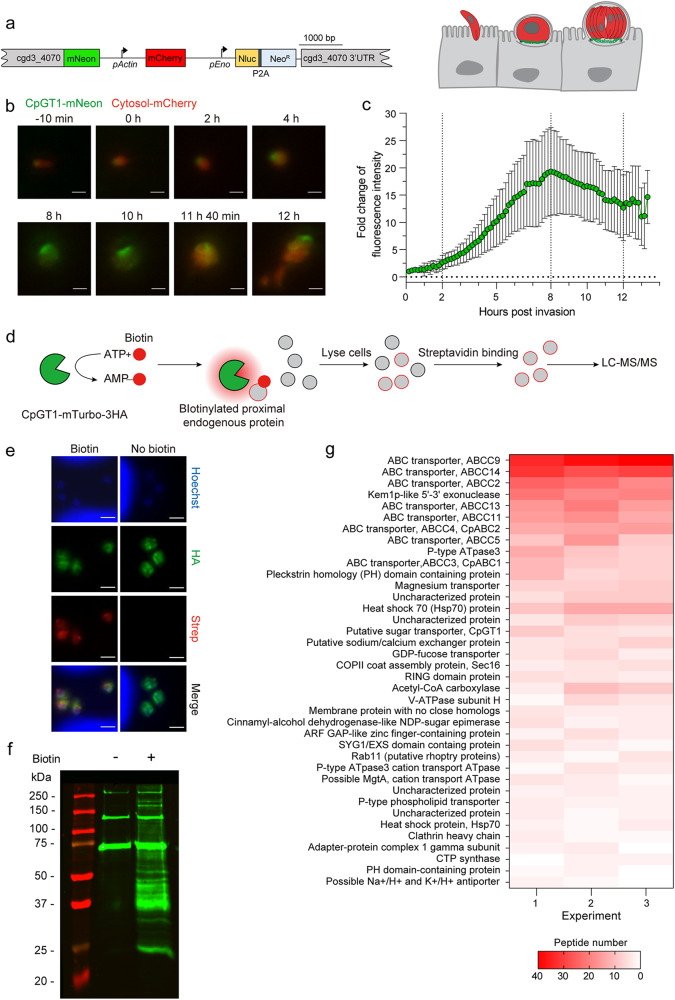
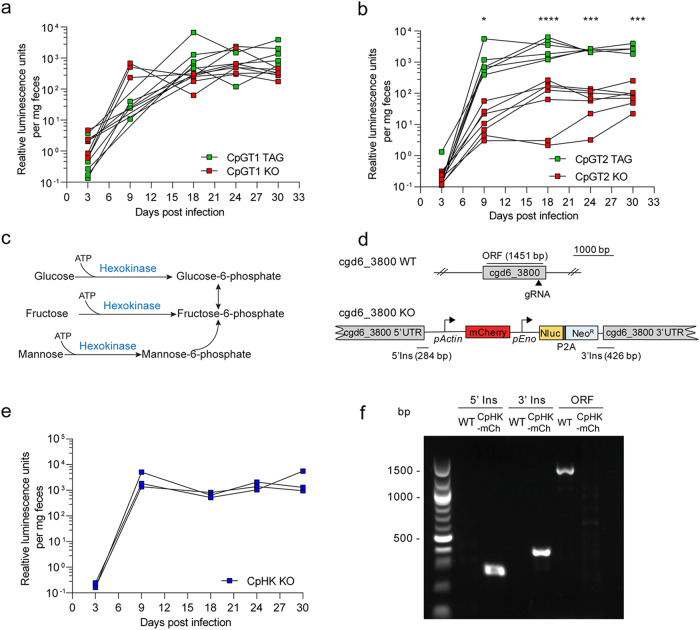
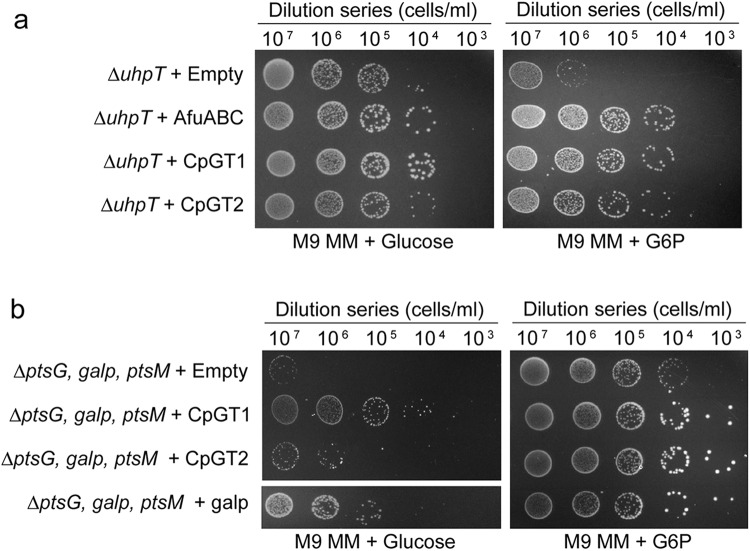
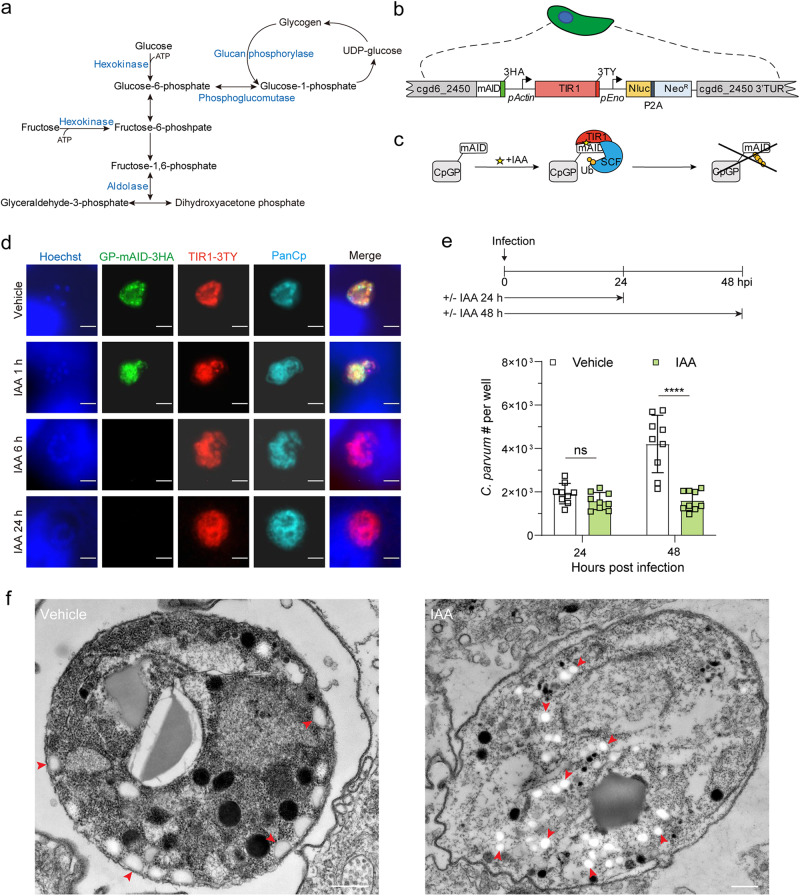
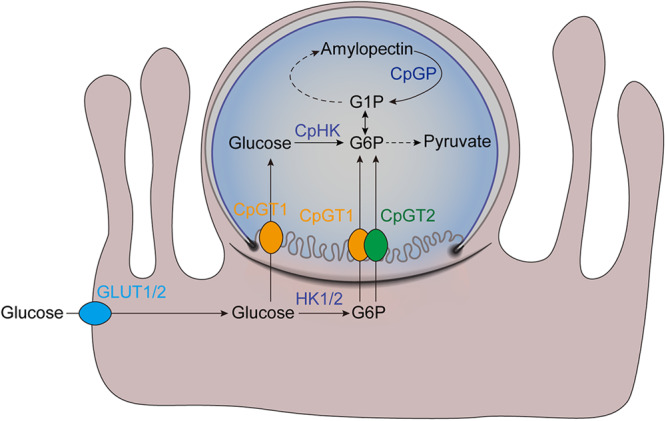
Cryptosporidium parvum is an obligate intracellular parasite with a highly reduced mitochondrion that lacks the tricarboxylic acid cycle and the ability to generate ATP, making the parasite reliant on glycolysis. Genetic ablation experiments demonstrated that neither of the two putative glucose transporters CpGT1 and CpGT2 were essential for growth. Surprisingly, hexokinase was also dispensable for parasite growth while the downstream enzyme aldolase was required, suggesting the parasite has an alternative way of obtaining phosphorylated hexose. Complementation studies in E. coli support a role for direct transport of glucose-6-phosphate from the host cell by the parasite transporters CpGT1 and CpGT2, thus bypassing a requirement for hexokinase. Additionally, the parasite obtains phosphorylated glucose from amylopectin stores that are released by the action of the essential enzyme glycogen phosphorylase. Collectively, these findings reveal that C. parvum relies on multiple pathways to obtain phosphorylated glucose both for glycolysis and to restore carbohydrate reserves.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Multiple pathways for glucose phosphate transport and utilization support growth of Cryptosporidium parvum.bioRxiv [Preprint]. 2023 Jun 27:2023.06.27.546703. doi: 10.1101/2023.06.27.546703. bioRxiv. 2023. Update in: Nat Commun. 2024 Jan 9;15(1):380. doi: 10.1038/s41467-024-44696-3. PMID: 37425855 Free PMC article. Updated. Preprint.
References
-
- Feng Y, Ryan UM, Xiao L. Genetic diversity and population structure of Cryptosporidium. Trends Parasitol. 2018;34:997–1011. - PubMed
-
- Ryan U, Fayer R, Xiao L. Cryptosporidium species in humans and animals: current understanding and research needs. Parasitology. 2014;141:1667–1685. - PubMed
-
- Kotloff KL, et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet. 2013;382:209–222. - PubMed
-
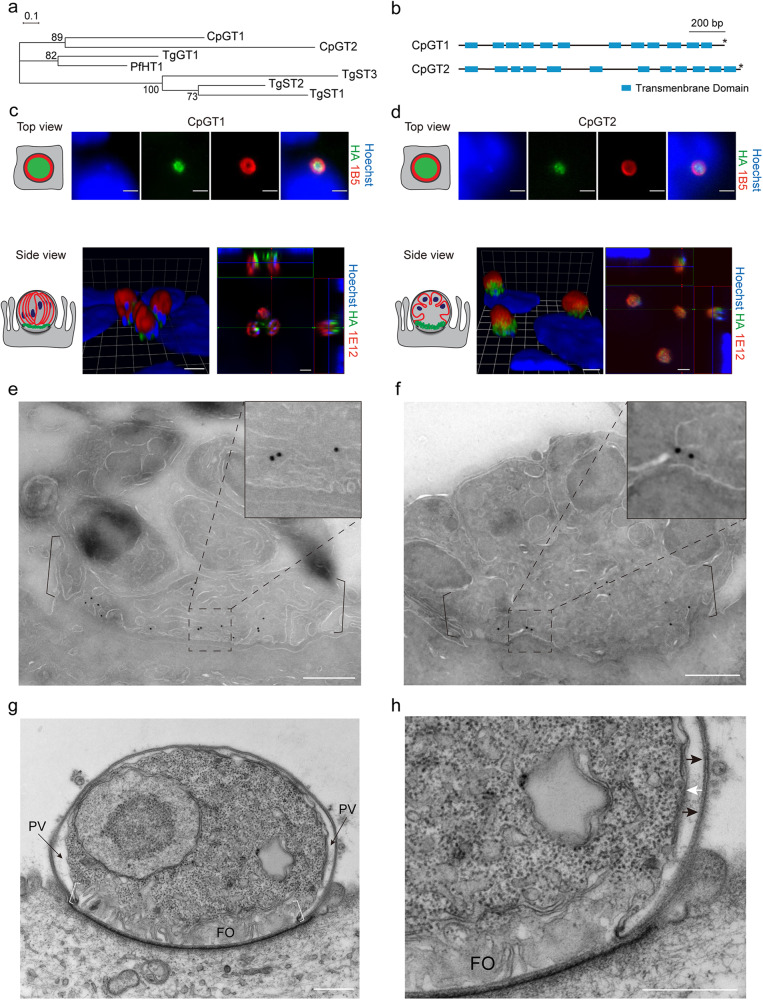
- Bochimoto H, Kondoh D, Ishihara Y, Kabir MHB, Kato K. Three-dimensional fine structure of feeder organelle in Cryptosporidium parvum. Parasitol. Int. 2019;73:101958. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
