

IFN-λ3 is induced by Leishmania donovani and can inhibit parasite growth in cell line models but not in the mouse model, while it shows a significant association with leishmaniasis in humans
- PMID: 38193711
- PMCID: PMC10863405
- DOI: 10.1128/iai.00504-23
IFN-λ3 is induced by Leishmania donovani and can inhibit parasite growth in cell line models but not in the mouse model, while it shows a significant association with leishmaniasis in humans
Abstract
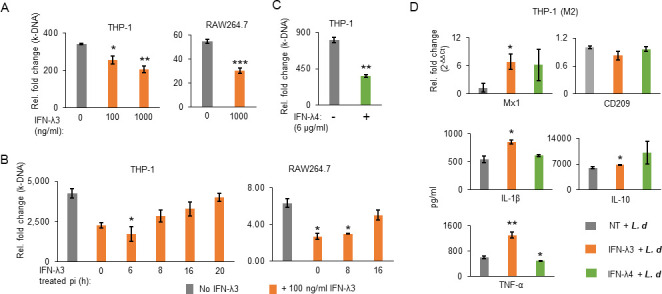
The intracellular protozoan parasite Leishmania donovani causes debilitating human diseases that involve visceral and dermal manifestations. Type 3 interferons (IFNs), also referred to as lambda IFNs (IFNL, IFN-L, or IFN-λ), are known to play protective roles against intracellular pathogens at the epithelial surfaces. Herein, we show that L. donovani induces IFN-λ3 in human as well as mouse cell line-derived macrophages. Interestingly, IFN-λ3 treatment significantly decreased parasite load in infected cells, mainly by increasing reactive oxygen species production. Microscopic examination showed that IFN-λ3 inhibited uptake but not replication, while the phagocytic ability of the cells was not affected. This was confirmed by experiments that showed that IFN-λ3 could decrease parasite load only when added to the medium at earlier time points, either during or soon after parasite uptake, but had no effect on parasite load when added at 24 h post-infection, suggesting that an early event during parasite uptake was targeted. Furthermore, the parasites could overcome the inhibitory effect of IFN-λ3, which was added at earlier time points, within 2-3 days post-infection. BALB/c mice treated with IFN-λ3 before infection led to a significant increase in expression of IL-4 and ARG1 post-infection in the spleen and liver, respectively, and to different pathological changes, especially in the liver, but not to changes in parasite load. Treatment with IFN-λ3 during infection did not decrease the parasite load in the spleen either. However, IFN-λ3 was significantly increased in the sera of visceral leishmaniasis patients, and the IFNL genetic variant rs12979860 was significantly associated with susceptibility to leishmaniasis.
Keywords: IFN-L3; Leishmania; Leishmania donovani; post-kala-azar dermal leishmaniasis; type III IFN; visceral leishmaniasis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








Similar articles
-
Leishmania eukaryotic initiation factor (LeIF) inhibits parasite growth in murine macrophages.PLoS One. 2014 May 15;9(5):e97319. doi: 10.1371/journal.pone.0097319. eCollection 2014. PLoS One. 2014. PMID: 24830439 Free PMC article.
-
Modulation of CD11C+ splenic dendritic cell functions in murine visceral leishmaniasis: correlation with parasite replication in the spleen.Immunology. 2000 Feb;99(2):305-13. doi: 10.1046/j.1365-2567.2000.00939.x. Immunology. 2000. PMID: 10692051 Free PMC article.
-
Tissue Specific Dual RNA-Seq Defines Host-Parasite Interplay in Murine Visceral Leishmaniasis Caused by Leishmania donovani and Leishmania infantum.Microbiol Spectr. 2022 Apr 27;10(2):e0067922. doi: 10.1128/spectrum.00679-22. Epub 2022 Apr 6. Microbiol Spectr. 2022. PMID: 35384718 Free PMC article.
-
Intracellular replication-deficient Leishmania donovani induces long lasting protective immunity against visceral leishmaniasis.J Immunol. 2009 Aug 1;183(3):1813-20. doi: 10.4049/jimmunol.0900276. Epub 2009 Jul 10. J Immunol. 2009. PMID: 19592661
-
The importance of T cell-derived cytokines in post-kala-azar dermal leishmaniasis.Cytokine. 2021 Nov;147:155321. doi: 10.1016/j.cyto.2020.155321. Epub 2020 Oct 8. Cytokine. 2021. PMID: 33039255 Review.
Cited by
-
Expression and function of interferon lambda receptor 1 variants.FEBS Lett. 2025 Feb;599(4):466-475. doi: 10.1002/1873-3468.15041. Epub 2024 Oct 22. FEBS Lett. 2025. PMID: 39435588 Review.
References
-
- Muniaraj M. 2014. The lost hope of elimination of kala-Azar (visceral Leishmaniasis) by 2010 and cyclic occurrence of its outbreak in India, blame falls on vector control practices or co-infection with human immunodeficiency virus or therapeutic modalities? Trop Parasitol 4:10–19. doi:10.4103/2229-5070.129143 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
