

This is a preprint.
Anti-Ebola virus mAb 3A6 with unprecedented potency protects highly viremic animals from fatal outcome and physically lifts its glycoprotein target from the virion membrane
- PMID: 38196595
- PMCID: PMC10775387
- DOI: 10.21203/rs.3.rs-3722563/v1
Anti-Ebola virus mAb 3A6 with unprecedented potency protects highly viremic animals from fatal outcome and physically lifts its glycoprotein target from the virion membrane
Update in
-
Anti-Ebola virus mAb 3A6 protects highly viremic animals from fatal outcome via binding GP(1,2) in a position elevated from the virion membrane.Nat Commun. 2025 Feb 3;16(1):1293. doi: 10.1038/s41467-025-56452-2. Nat Commun. 2025. PMID: 39900911 Free PMC article.
Abstract
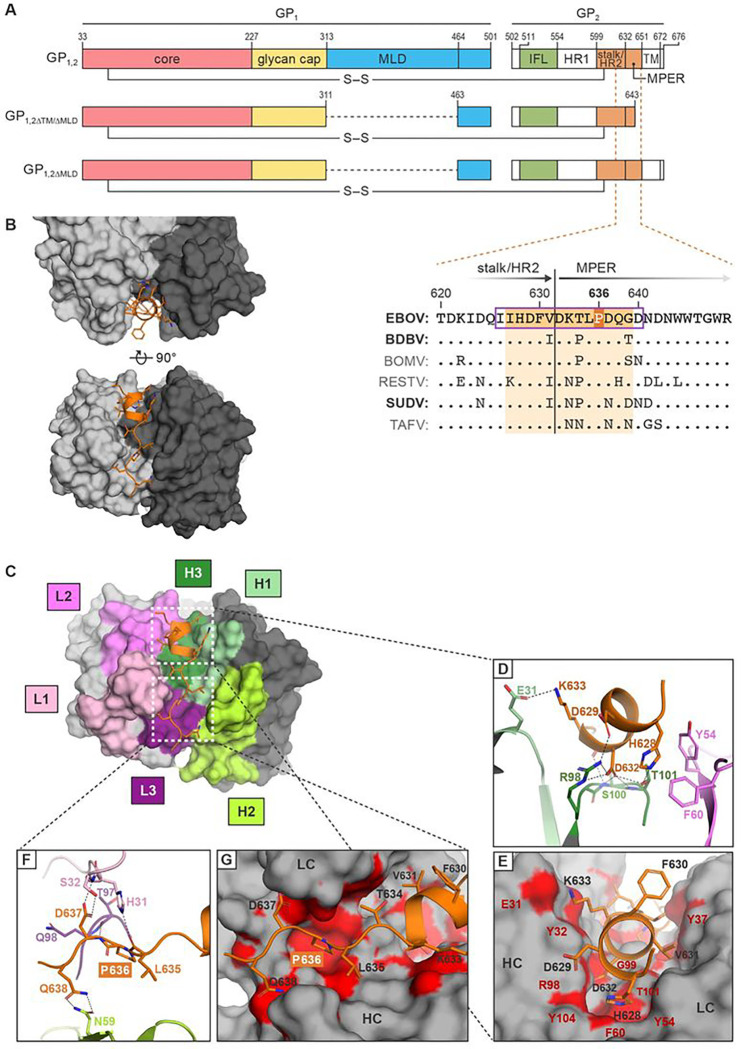
Monoclonal antibodies (mAbs) against Ebola virus (EBOV) glycoprotein (GP1,2) are the standard of care for Ebola virus disease (EVD). Anti-GP1,2 mAbs targeting the stalk and membrane proximal external region (MPER) potently neutralize EBOV in vitro. However, their neutralization mechanism is poorly understood because they target a GP1,2 epitope that has evaded structural characterization. Moreover, their in vivo efficacy has only been evaluated in the mouse model of EVD. Using x-ray crystallography and cryo-electron tomography of 3A6 complexed with its stalk- GP1,2 MPER epitope we reveal a novel mechanism in which 3A6 elevates the stalk or stabilizes a conformation of GP1,2 that is lifted from the virion membrane. In domestic guinea pig and rhesus monkey EVD models, 3A6 provides therapeutic benefit at high viremia levels, advanced disease stages, and at the lowest dose yet demonstrated for any anti-EBOV mAb-based monotherapy. These findings can guide design of next-generation, highly potent anti-EBOV mAbs.
Keywords: 3A6; Animal study; EBOV; Ebola virus; MPER; antibody; efficacy study; mAb; membrane proximal external region; monoclonal antibody; stalk.
Conflict of interest statement
Competing interests A.G., E.D., and B.J.D. are employees of Integral Molecular, and B.J.D.is a shareholder in that company. Additional Declarations: There is NO Competing Interest.
Figures




References
-
- Kuhn J.H., Amarasinghe G.K. & Perry D.L. Filoviridae. in Fields Virology Vol. 1 (Emerging Viruses) (eds. Howley P.M., Knipe D.M. & Whelan S.P.J.) 449–503 (Wolters Kluwer/Lippincott Williams & Wilkins, Philadelphia, Pennsylvania, USA, 2020).
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
