

Silencing dentate newborn neurons alters excitatory/inhibitory balance and impairs behavioral inhibition and flexibility
- PMID: 38198539
- PMCID: PMC10780870
- DOI: 10.1126/sciadv.adk4741
Silencing dentate newborn neurons alters excitatory/inhibitory balance and impairs behavioral inhibition and flexibility
Abstract
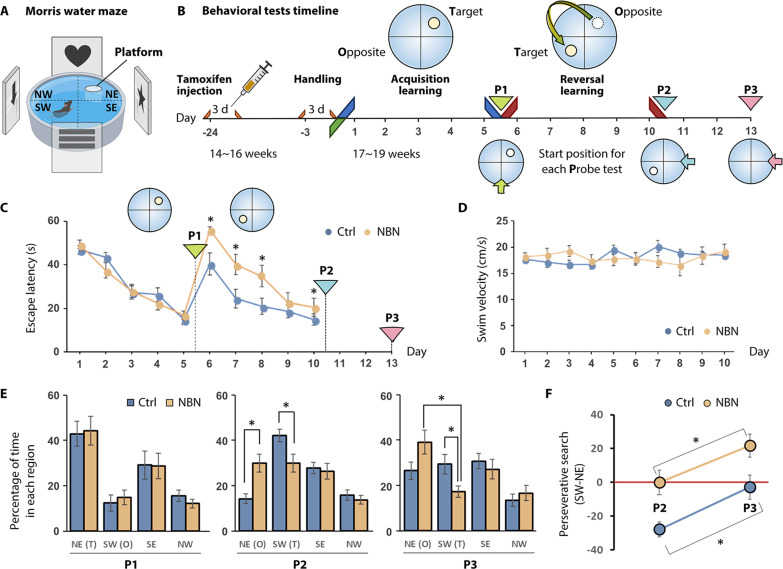
Adult neurogenesis confers the hippocampus with unparalleled neural plasticity, essential for intricate cognitive functions. The specific influence of sparse newborn neurons (NBNs) in modulating neural activities and subsequently steering behavior, however, remains obscure. Using an engineered NBN-tetanus toxin mouse model (NBN-TeTX), we noninvasively silenced NBNs, elucidating their crucial role in impulse inhibition and cognitive flexibility as evidenced through Morris water maze reversal learning and Go/Nogo task in operant learning. Task-based functional MRI (tb-fMRI) paired with operant learning revealed dorsal hippocampal hyperactivation during the Nogo task in male NBN-TeTX mice, suggesting that hippocampal hyperexcitability might underlie the observed behavioral deficits. Additionally, resting-state fMRI (rs-fMRI) exhibited enhanced functional connectivity between the dorsal and ventral dentate gyrus following NBN silencing. Further investigations into the activities of PV+ interneurons and mossy cells highlighted the indispensability of NBNs in maintaining the hippocampal excitation/inhibition balance. Our findings emphasize that the neural plasticity driven by NBNs extensively modulates the hippocampus, sculpting inhibitory control and cognitive flexibility.
Figures







References
-
- Ryan T. J., Frankland P. W., Forgetting as a form of adaptive engram cell plasticity. Nat. Rev. Neurosci. 23, 173–186 (2022). - PubMed
-
- Miyake A., Friedman N. P., Emerson M. J., Witzki A. H., Howerter A., Wager T. D., The unity and diversity of executive functions and their contributions to complex “Frontal Lobe” tasks: A latent variable analysis. Cogn. Psychol. 41, 49–100 (2000). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
