

Adding α,α-disubstituted and β-linked monomers to the genetic code of an organism
- PMID: 38200312
- PMCID: PMC10794150
- DOI: 10.1038/s41586-023-06897-6
Adding α,α-disubstituted and β-linked monomers to the genetic code of an organism
Abstract
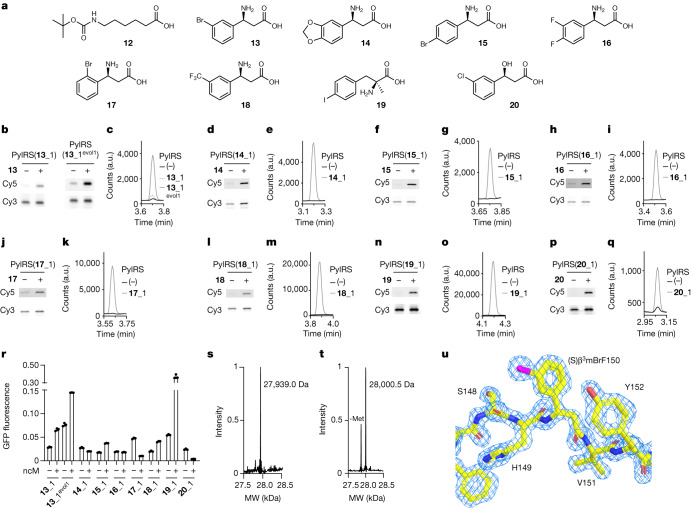
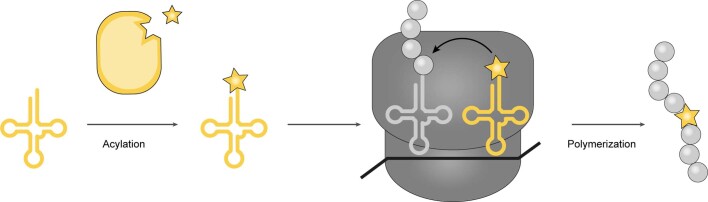
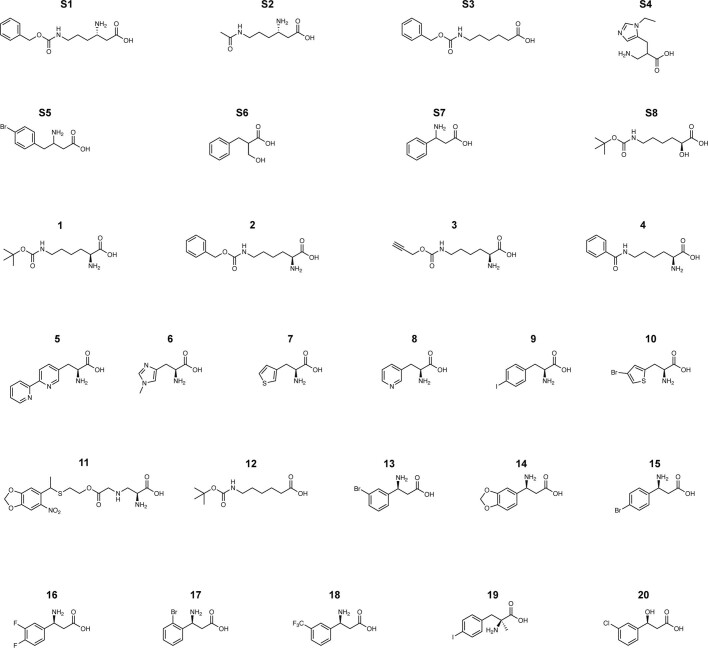
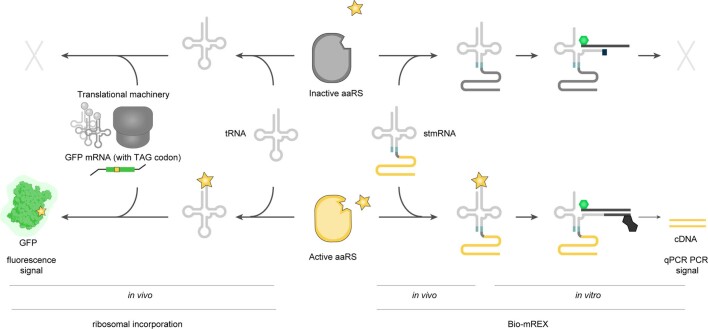
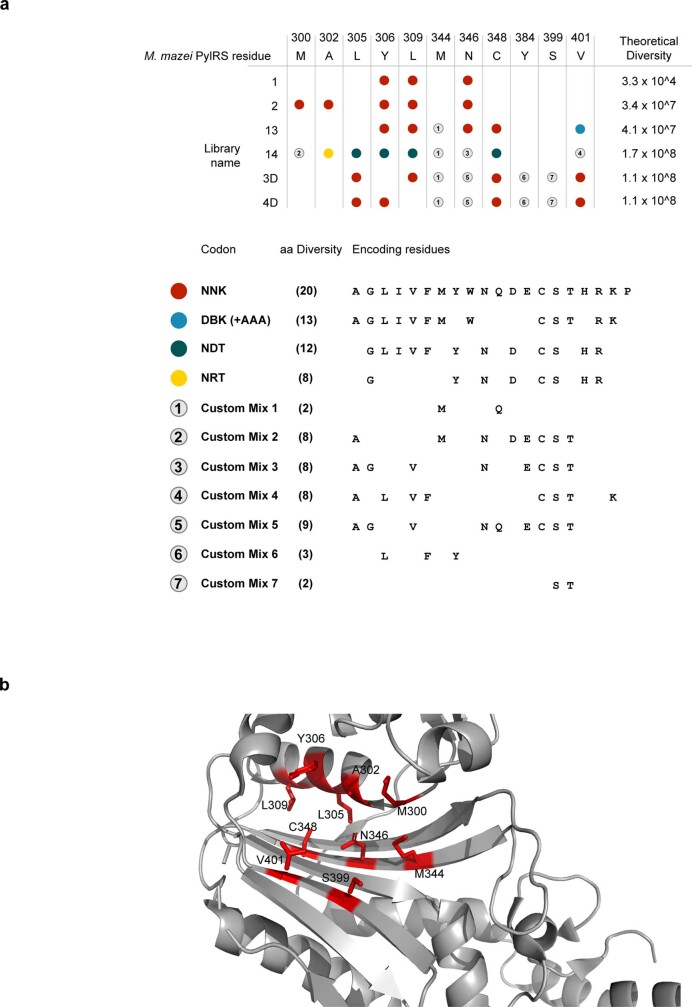
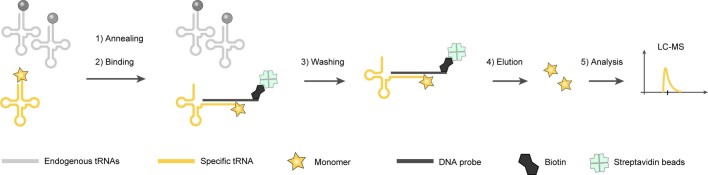
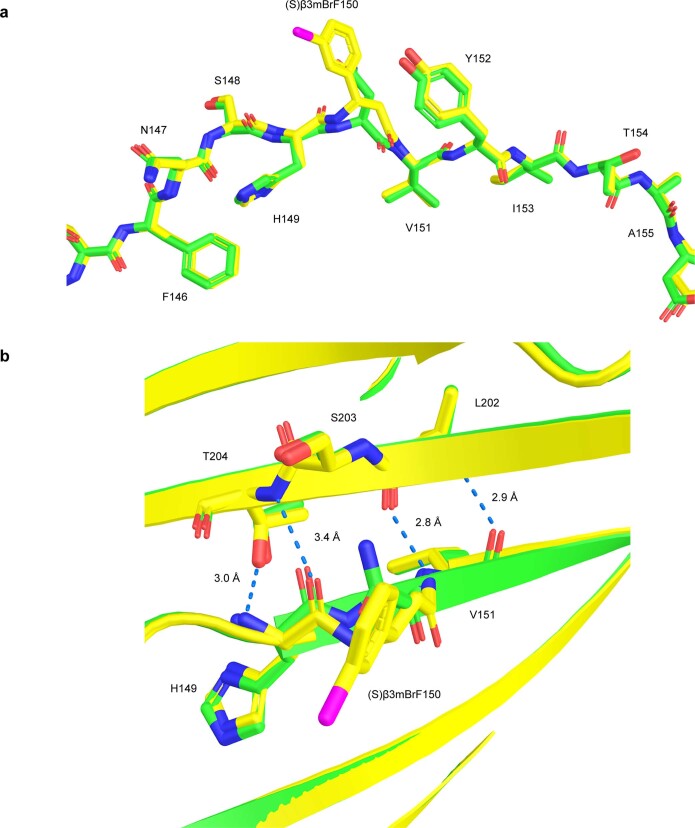
The genetic code of living cells has been reprogrammed to enable the site-specific incorporation of hundreds of non-canonical amino acids into proteins, and the encoded synthesis of non-canonical polymers and macrocyclic peptides and depsipeptides1-3. Current methods for engineering orthogonal aminoacyl-tRNA synthetases to acylate new monomers, as required for the expansion and reprogramming of the genetic code, rely on translational readouts and therefore require the monomers to be ribosomal substrates4-6. Orthogonal synthetases cannot be evolved to acylate orthogonal tRNAs with non-canonical monomers (ncMs) that are poor ribosomal substrates, and ribosomes cannot be evolved to polymerize ncMs that cannot be acylated onto orthogonal tRNAs-this co-dependence creates an evolutionary deadlock that has essentially restricted the scope of translation in living cells to α-L-amino acids and closely related hydroxy acids. Here we break this deadlock by developing tRNA display, which enables direct, rapid and scalable selection for orthogonal synthetases that selectively acylate their cognate orthogonal tRNAs with ncMs in Escherichia coli, independent of whether the ncMs are ribosomal substrates. Using tRNA display, we directly select orthogonal synthetases that specifically acylate their cognate orthogonal tRNA with eight non-canonical amino acids and eight ncMs, including several β-amino acids, α,α-disubstituted-amino acids and β-hydroxy acids. We build on these advances to demonstrate the genetically encoded, site-specific cellular incorporation of β-amino acids and α,α-disubstituted amino acids into a protein, and thereby expand the chemical scope of the genetic code to new classes of monomers.
© 2024. The Author(s).
Conflict of interest statement
J.W.C. is a founder of Constructive Bio. The Medical Research Council have filed a patent application based on this work. The other authors declare no competing interests.
Figures
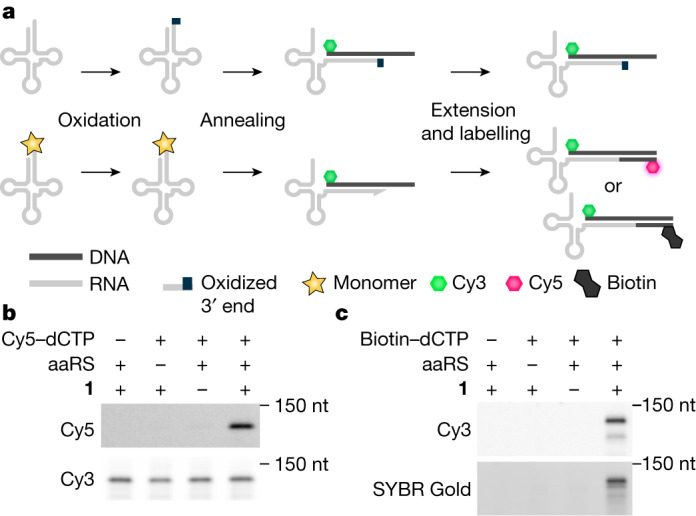
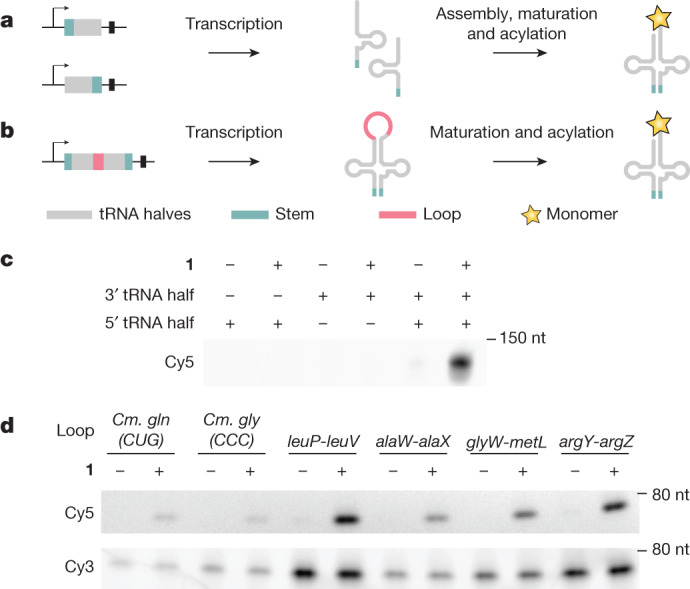
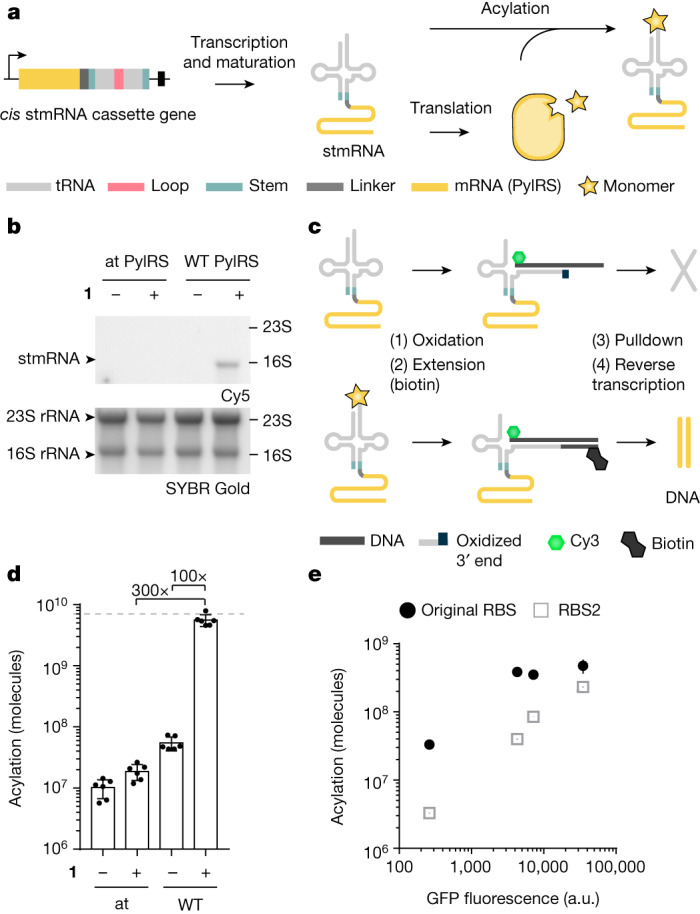
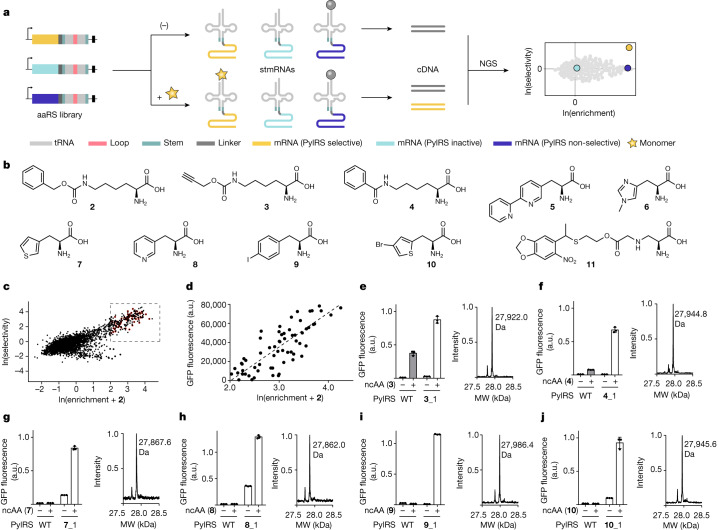
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
