

Thymic Carcinoma: Unraveling Neuroendocrine Differentiation and Epithelial Cell Identity Loss
- PMID: 38201543
- PMCID: PMC10778300
- DOI: 10.3390/cancers16010115
Thymic Carcinoma: Unraveling Neuroendocrine Differentiation and Epithelial Cell Identity Loss
Abstract
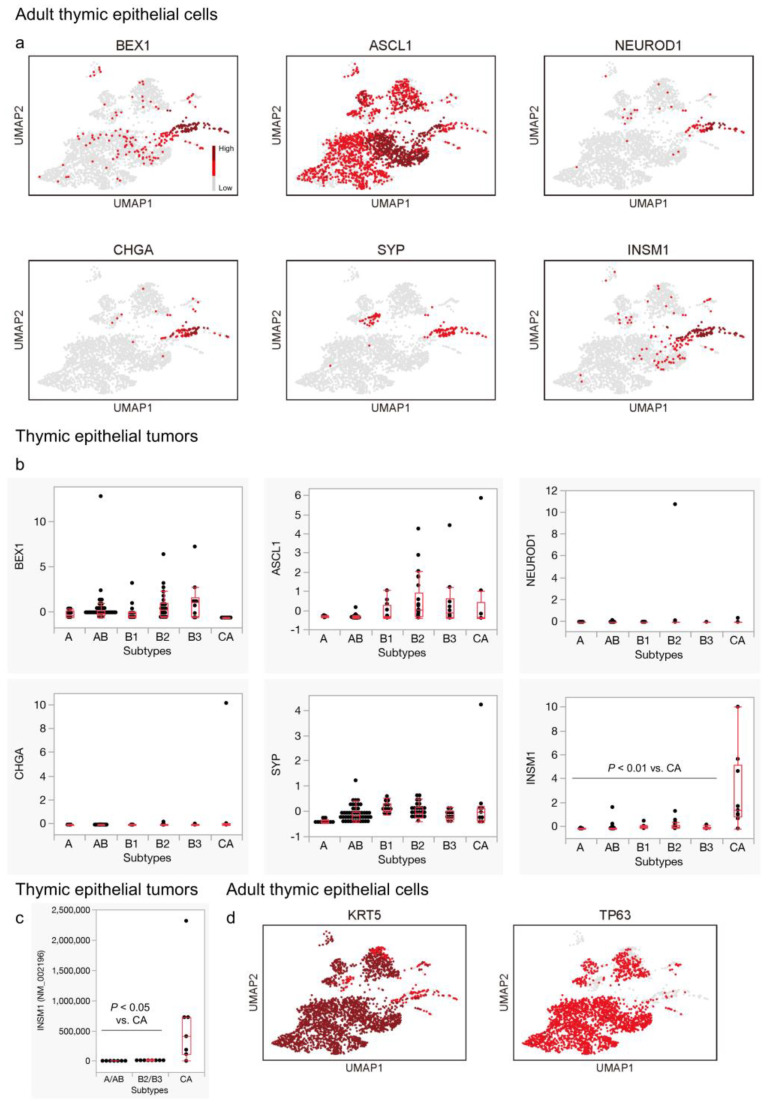
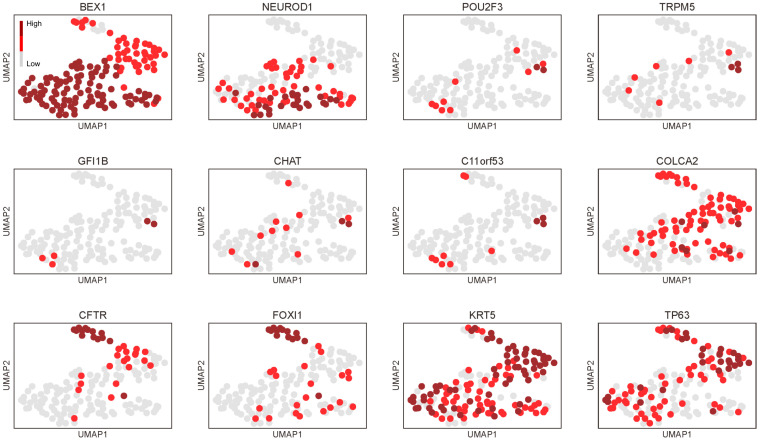
Background: The histogenesis of thymic epithelial tumors (TETs) has been a subject of debate. Recent technological advancements have revealed that thymic carcinomas often exhibit a phenotype akin to tuft cells, which is a subset of medullary TECs. Here, we further explored the gene expression signatures of thymic carcinomas in relation to tuft cells and their kinships-ionocytes and neuroendocrine cells (neuroendocrine group).
Methods: We analyzed a single-cell RNA sequencing dataset from the normal human thymus. Concurrently, we examined publicly available datasets on the mRNA expression and methylation status of TECs and lung cancers. Real-time quantitative PCR was also conducted with our tissue samples.
Results: Thymic carcinomas displayed a neuroendocrine phenotype biased toward tuft cells and ionocytes. When exploring the possible regulators of this phenotype, we discovered that HDAC9 and NFATC1 were characteristically expressed in the neuroendocrine group in adult TECs and thymic carcinomas. Additionally, the pan-thymic epithelium markers, exemplified by PAX9 and SIX1, were significantly suppressed in thymic carcinomas.
Conclusions: Thymic carcinomas might be characterized by unique neuroendocrine differentiation and loss of identity as thymic epithelial cells. Future studies investigating the role of HDAC9 and NFATC1 in thymic epithelium are warranted to explore their potential as therapeutic targets in TETs.
Keywords: HDAC9; NFATC1; PAX9; SIX1; ionocytes; neuroendocrine cells; thymic carcinoma; thymic epithelial tumors; thymoma; tuft cells.
Conflict of interest statement
The authors declare that they have no competing interest.
Figures






Similar articles
-
A Tuft Cell-Like Signature Is Highly Prevalent in Thymic Squamous Cell Carcinoma and Delineates New Molecular Subsets Among the Major Lung Cancer Histotypes.J Thorac Oncol. 2021 Jun;16(6):1003-1016. doi: 10.1016/j.jtho.2021.02.008. Epub 2021 Feb 17. J Thorac Oncol. 2021. PMID: 33609752
-
Histogenetic and disease-relevant phenotypes in thymic epithelial tumors (TETs): The potential significance for future TET classification.Pathol Int. 2023 Jul;73(7):265-280. doi: 10.1111/pin.13343. Epub 2023 Jun 6. Pathol Int. 2023. PMID: 37278579 Review.
-
Cytokeratins in normal thymus and thymic epithelial tumors.Cancer. 1993 Jan 1;71(1):99-105. doi: 10.1002/1097-0142(19930101)71:1<99::aid-cncr2820710116>3.0.co;2-6. Cancer. 1993. PMID: 7677978
-
CD90 Marks a Mesenchymal Program in Human Thymic Epithelial Cells In Vitro and In Vivo.Front Immunol. 2022 Mar 16;13:846281. doi: 10.3389/fimmu.2022.846281. eCollection 2022. Front Immunol. 2022. PMID: 35371075 Free PMC article.
-
Molecular landscape of thymic epithelial tumors.Semin Diagn Pathol. 2022 Mar;39(2):131-136. doi: 10.1053/j.semdp.2021.06.011. Epub 2021 Jul 3. Semin Diagn Pathol. 2022. PMID: 34272124 Review.
Cited by
-
Thymic Mimetic Cells: Evolutionarily Ancient Mirrors of the Periphery.Immunol Rev. 2025 May;331(1):e70028. doi: 10.1111/imr.70028. Immunol Rev. 2025. PMID: 40247628 Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
