

3'dNTP Binding Is Modulated during Primer Synthesis and Translesion by Human PrimPol
- PMID: 38203225
- PMCID: PMC10778844
- DOI: 10.3390/ijms25010051
3'dNTP Binding Is Modulated during Primer Synthesis and Translesion by Human PrimPol
Abstract
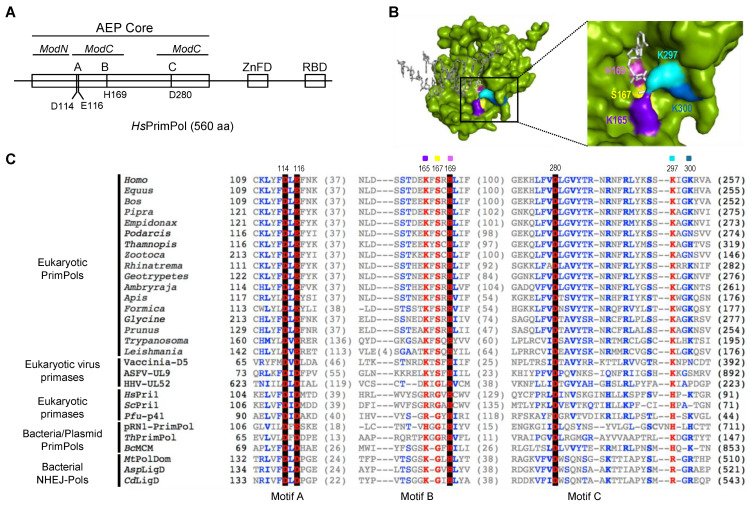
PrimPol is a DNA primase/polymerase from the Archaeo-Eukaryotic Primase (AEP) superfamily that enables the progression of stalled replication forks by synthesizing DNA primers ahead of blocking lesions or abnormal structures in the ssDNA template. PrimPol's active site is formed by three AEP-conserved motifs: A, B and C. Motifs A and C of human PrimPol (HsPrimPol) harbor the catalytic residues (Asp114, Glu116, Asp280) acting as metal ligands, whereas motif B includes highly conserved residues (Lys165, Ser167 and His169), which are postulated to stabilize 3' incoming deoxynucleotides (dNTPs). Additionally, other putative nucleotide ligands are situated close to motif C: Lys297, almost invariant in the whole AEP superfamily, and Lys300, specifically conserved in eukaryotic PrimPols. Here, we demonstrate that His169 is absolutely essential for 3'dNTP binding and, hence, for both primase and polymerase activities of HsPrimPol, whereas Ser167 and Lys297 are crucial for the dimer synthesis initiation step during priming, but dispensable for subsequent dNTP incorporation on growing primers. Conversely, the elimination of Lys165 does not affect the overall primase function; however, it is required for damage avoidance via primer-template realignments. Finally, Lys300 is identified as an extra anchor residue to stabilize the 3' incoming dNTP. Collectively, these results demonstrate that individual ligands modulate the stabilization of 3' incoming dNTPs to optimize DNA primer synthesis efficiency during initiation and primer maturation.
Keywords: 3′site dNTP ligands; AEP; AEP motif B; TLS; human PrimPol.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
Motif WFYY of human PrimPol is crucial to stabilize the incoming 3'-nucleotide during replication fork restart.Nucleic Acids Res. 2021 Aug 20;49(14):8199-8213. doi: 10.1093/nar/gkab634. Nucleic Acids Res. 2021. PMID: 34302490 Free PMC article.
-
The invariant glutamate of human PrimPol DxE motif is critical for its Mn2+-dependent distinctive activities.DNA Repair (Amst). 2019 May;77:65-75. doi: 10.1016/j.dnarep.2019.03.006. Epub 2019 Mar 14. DNA Repair (Amst). 2019. PMID: 30889508
-
Mechanism of DNA primer synthesis by human PrimPol.Enzymes. 2019;45:289-310. doi: 10.1016/bs.enz.2019.06.003. Epub 2019 Jul 13. Enzymes. 2019. PMID: 31627881 Review.
-
Human PrimPol Discrimination against Dideoxynucleotides during Primer Synthesis.Genes (Basel). 2021 Sep 24;12(10):1487. doi: 10.3390/genes12101487. Genes (Basel). 2021. PMID: 34680882 Free PMC article.
-
PrimPol: A Breakthrough among DNA Replication Enzymes and a Potential New Target for Cancer Therapy.Biomolecules. 2022 Feb 3;12(2):248. doi: 10.3390/biom12020248. Biomolecules. 2022. PMID: 35204749 Free PMC article. Review.
Cited by
-
Mouse PrimPol Outperforms Its Human Counterpart as a Robust DNA Primase.Int J Mol Sci. 2025 Jul 19;26(14):6947. doi: 10.3390/ijms26146947. Int J Mol Sci. 2025. PMID: 40725194 Free PMC article.
-
A four-in-one replicase integrating key enzymatic activities for DNA replication.Nucleic Acids Res. 2025 Jun 20;53(12):gkaf542. doi: 10.1093/nar/gkaf542. Nucleic Acids Res. 2025. PMID: 40548937 Free PMC article.
References
-
- Bianchi J., Rudd S.G., Jozwiakowski S.K., Bailey L.J., Soura V., Taylor E., Stevanovic I., Green A.J., Stracker T.H., Lindsay H.D., et al. PrimPol Bypasses UV Photoproducts during Eukaryotic Chromosomal DNA Replication. Mol. Cell. 2013;52:566–573. doi: 10.1016/j.molcel.2013.10.035. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
