

Hexavalent Chromium Targets Securin to Drive Numerical Chromosome Instability in Human Lung Cells
- PMID: 38203427
- PMCID: PMC10778806
- DOI: 10.3390/ijms25010256
Hexavalent Chromium Targets Securin to Drive Numerical Chromosome Instability in Human Lung Cells
Abstract
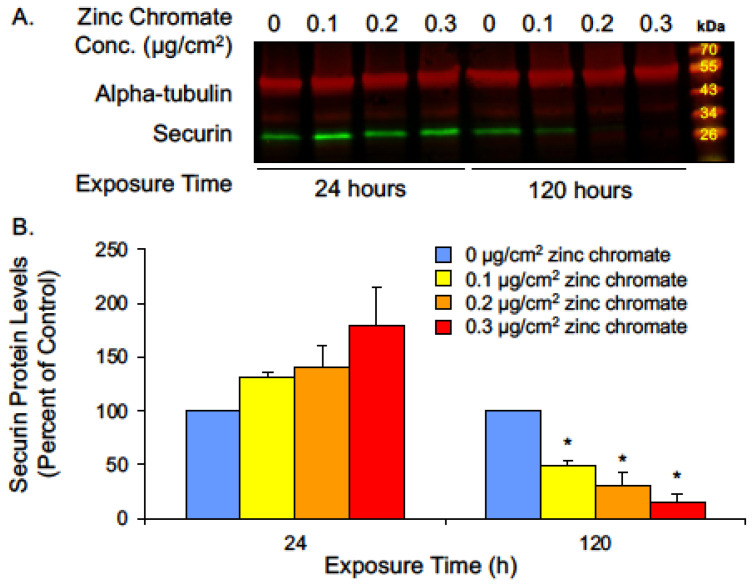
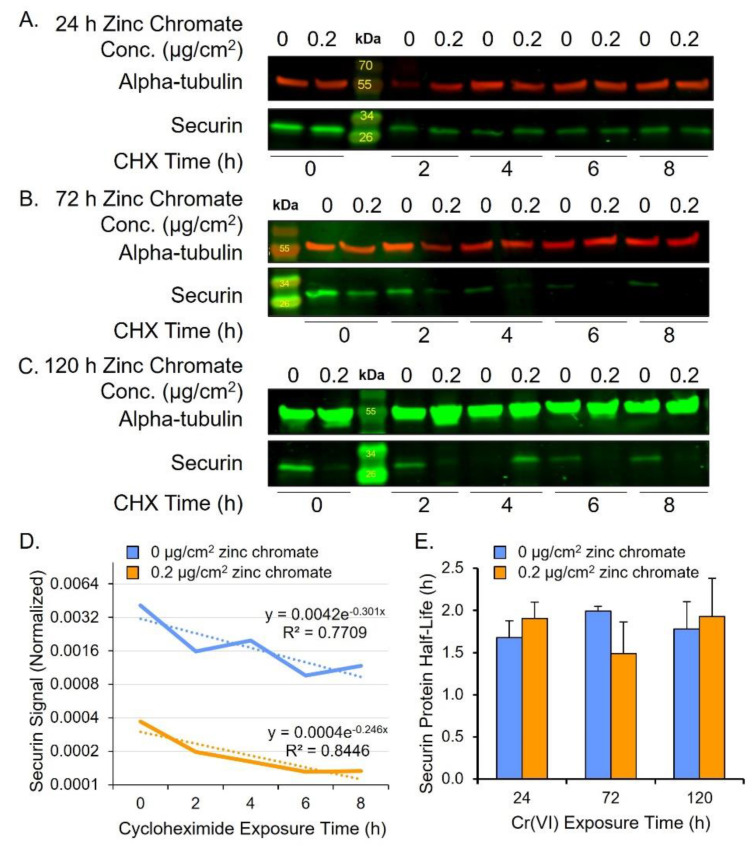
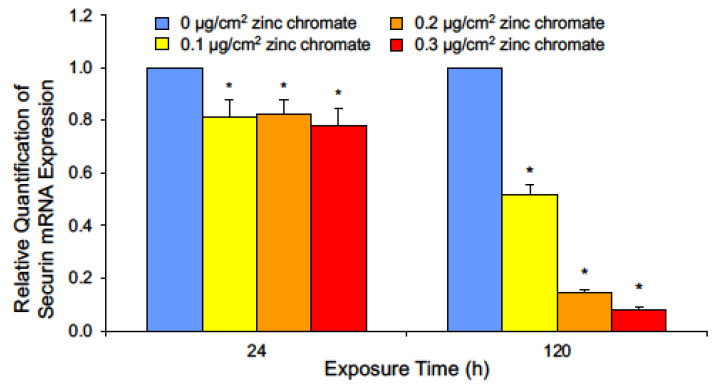
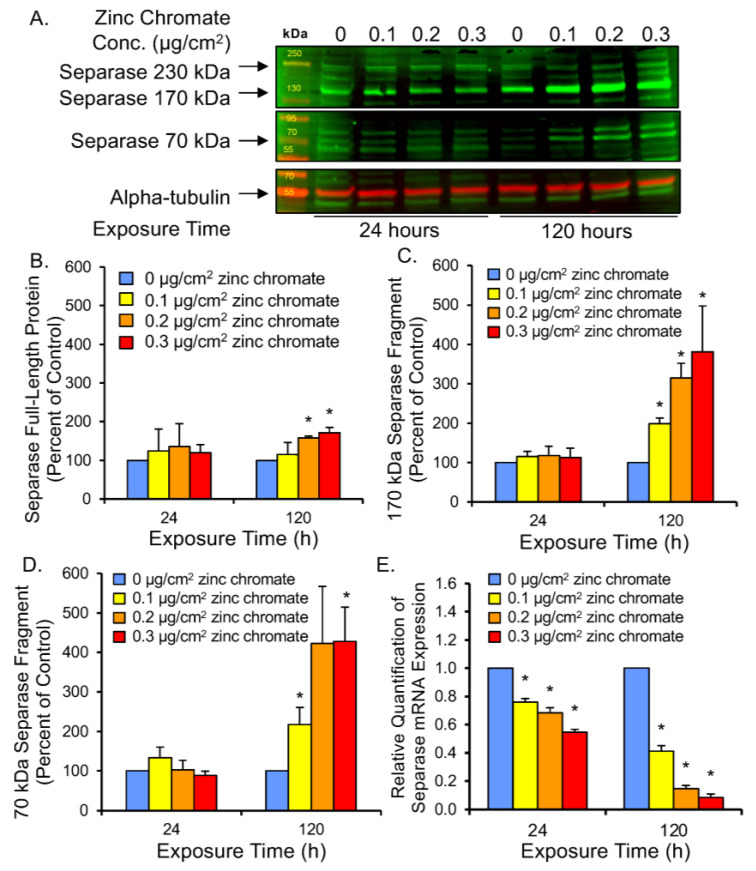
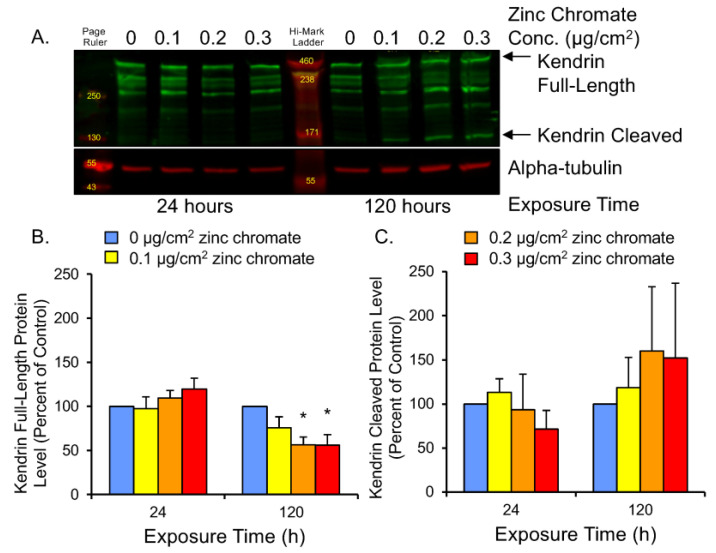
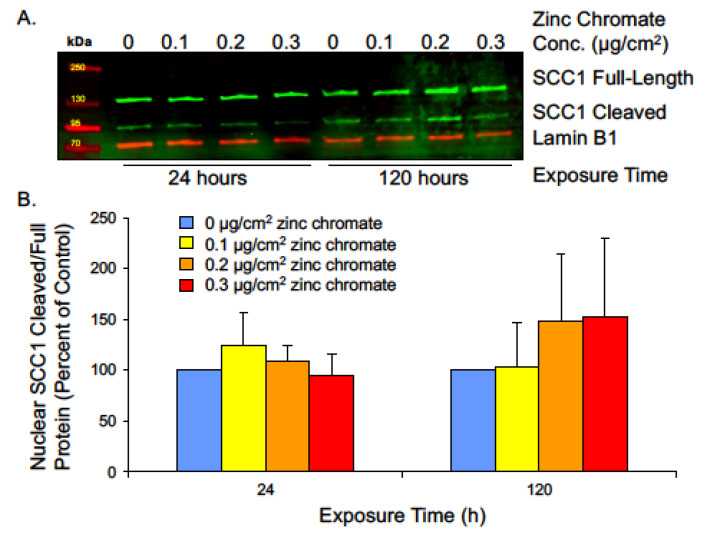
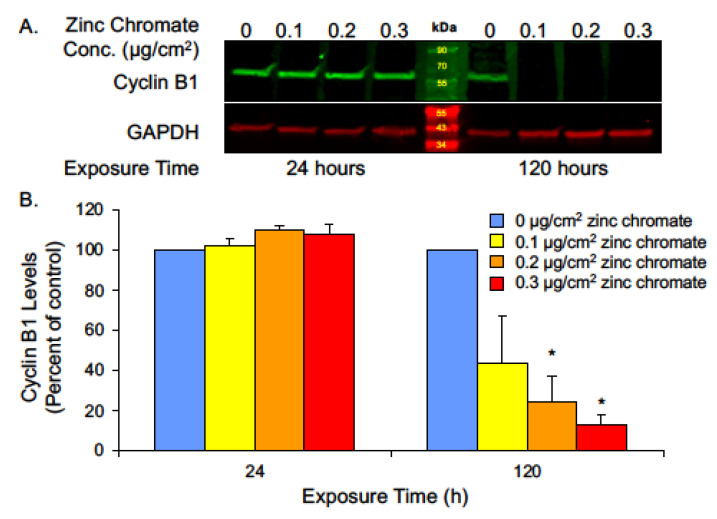
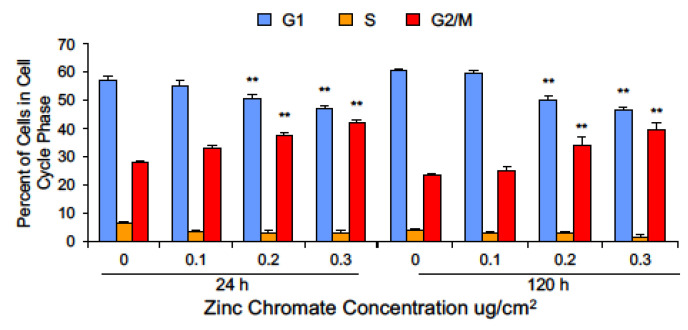
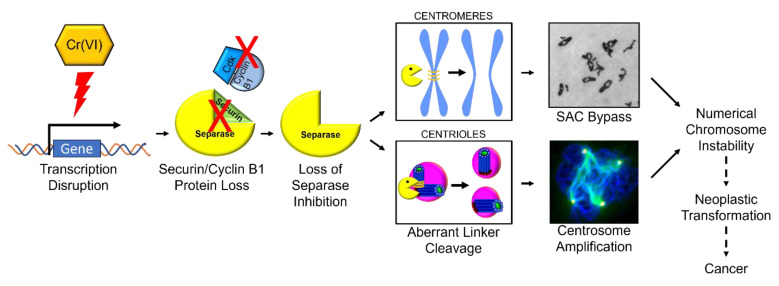
Hexavalent chromium [Cr(VI)] is a known human lung carcinogen with widespread exposure in environmental and occupational settings. Despite well-known cancer risks, the molecular mechanisms of Cr(VI)-induced carcinogenesis are not well understood, but a major driver of Cr(VI) carcinogenesis is chromosome instability. Previously, we reported Cr(VI) induced numerical chromosome instability, premature centriole disengagement, centrosome amplification, premature centromere division, and spindle assembly checkpoint bypass. A key regulator of these events is securin, which acts by regulating the cleavage ability of separase. Thus, in this study we investigated securin disruption by Cr(VI) exposure. We exposed human lung cells to a particulate Cr(VI) compound, zinc chromate, for acute (24 h) and prolonged (120 h) time points. We found prolonged Cr(VI) exposure caused marked decrease in securin levels and function. After prolonged exposure at the highest concentration, securin protein levels were decreased to 15.3% of control cells, while securin mRNA quantification was 7.9% relative to control cells. Additionally, loss of securin function led to increased separase activity manifested as enhanced cleavage of separase substrates; separase, kendrin, and SCC1. These data show securin is targeted by prolonged Cr(VI) exposure in human lung cells. Thus, a new mechanistic model for Cr(VI)-induced carcinogenesis emerges with centrosome and centromere disruption as key components of numerical chromosome instability, a key driver in Cr(VI) carcinogenesis.
Keywords: centrosome amplification; chromosome instability; hexavalent chromium; securin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Carcinogenic Mechanisms of Hexavalent Chromium: From DNA Breaks to Chromosome Instability and Neoplastic Transformation.Curr Environ Health Rep. 2024 Dec;11(4):484-546. doi: 10.1007/s40572-024-00460-9. Epub 2024 Oct 28. Curr Environ Health Rep. 2024. PMID: 39466546 Review.
-
A whale of a tale: whale cells evade the driving mechanism for hexavalent chromium-induced chromosome instability.Toxicol Sci. 2024 Apr 29;199(1):49-62. doi: 10.1093/toxsci/kfae030. Toxicol Sci. 2024. PMID: 38539048 Free PMC article.
-
Chronic Exposure to Particulate Chromate Induces Premature Centrosome Separation and Centriole Disengagement in Human Lung Cells.Toxicol Sci. 2015 Oct;147(2):490-9. doi: 10.1093/toxsci/kfv146. Epub 2015 Aug 19. Toxicol Sci. 2015. PMID: 26293554 Free PMC article.
-
Chronic exposure to particulate chromate induces spindle assembly checkpoint bypass in human lung cells.Chem Res Toxicol. 2006 Nov;19(11):1492-8. doi: 10.1021/tx0601410. Chem Res Toxicol. 2006. PMID: 17112237
-
Structure and Function of the Separase-Securin Complex.Subcell Biochem. 2021;96:217-232. doi: 10.1007/978-3-030-58971-4_4. Subcell Biochem. 2021. PMID: 33252730 Free PMC article. Review.
Cited by
-
Carcinogenic Mechanisms of Hexavalent Chromium: From DNA Breaks to Chromosome Instability and Neoplastic Transformation.Curr Environ Health Rep. 2024 Dec;11(4):484-546. doi: 10.1007/s40572-024-00460-9. Epub 2024 Oct 28. Curr Environ Health Rep. 2024. PMID: 39466546 Review.
-
A whale of a tale: whale cells evade the driving mechanism for hexavalent chromium-induced chromosome instability.Toxicol Sci. 2024 Apr 29;199(1):49-62. doi: 10.1093/toxsci/kfae030. Toxicol Sci. 2024. PMID: 38539048 Free PMC article.
-
Particulate hexavalent chromium inhibits global transcription of genes in DNA repair pathways, particularly targeting homologous recombination repair, base excision repair, mismatch repair and microhomology-mediated end-joining.J Hazard Mater. 2025 Mar 5;485:136892. doi: 10.1016/j.jhazmat.2024.136892. Epub 2024 Dec 15. J Hazard Mater. 2025. PMID: 39706010
-
Transcriptomic analysis reveals particulate hexavalent chromium regulates key inflammatory pathways in human lung fibroblasts as a possible mechanism of carcinogenesis.Toxicol Appl Pharmacol. 2024 Apr;485:116889. doi: 10.1016/j.taap.2024.116889. Epub 2024 Mar 11. Toxicol Appl Pharmacol. 2024. PMID: 38479592 Free PMC article.
References
-
- Wilbur S., Abadin H., Fay M., Yu D., Tencza B., Ingerman L., Klotzbach J., James S. Toxicological Profile for Chromium. Agency for Toxic Substances and Disease Registry (ATSDR) Toxicological Profiles; Agency for Toxic Substances and Disease Registry (US); Atlanta, GA, USA: 2012. - PubMed
-
- IARC Working Group on the Evaluation of Carcinogenic Risk to Humans . Arsenic, Metals, Fibres and Dusts. International Agency for Research on Cancer; Lyon, France: 2012.
-
- Langard S. Role of Chemical Species and Exposure Characteristics in Cancer among Persons Occupationally Exposed to Chromium Compounds. Scand. J. Work. Environ. Health. 1993;19((Suppl. 1)):81–89. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
