

Astrocytic Regulation of Endocannabinoid-Dependent Synaptic Plasticity in the Dorsolateral Striatum
- PMID: 38203752
- PMCID: PMC10779090
- DOI: 10.3390/ijms25010581
Astrocytic Regulation of Endocannabinoid-Dependent Synaptic Plasticity in the Dorsolateral Striatum
Abstract
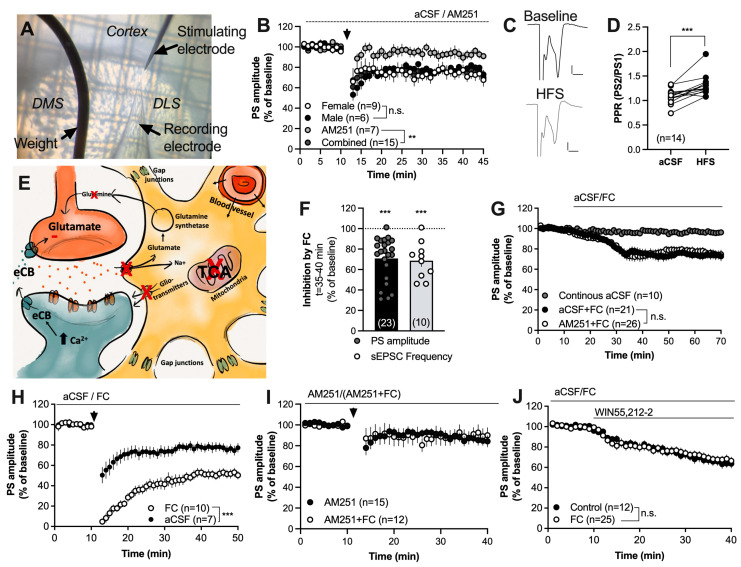
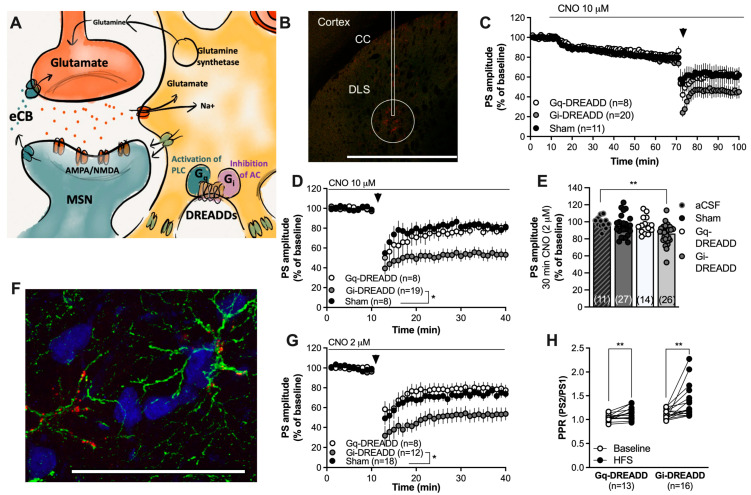
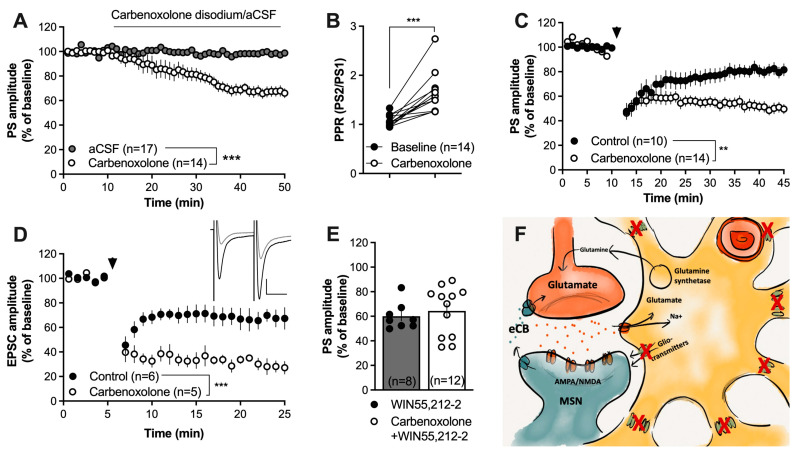
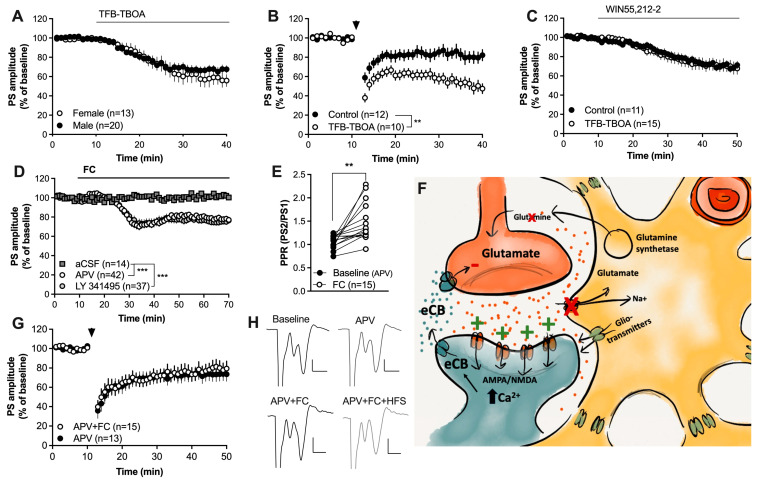
Astrocytes are pivotal for synaptic transmission and may also play a role in the induction and expression of synaptic plasticity, including endocannabinoid-mediated long-term depression (eCB-LTD). In the dorsolateral striatum (DLS), eCB signaling plays a major role in balancing excitation and inhibition and promoting habitual learning. The aim of this study was to outline the role of astrocytes in regulating eCB signaling in the DLS. To this end, we employed electrophysiological slice recordings combined with metabolic, chemogenetic and pharmacological approaches in an attempt to selectively suppress astrocyte function. High-frequency stimulation induced eCB-mediated LTD (HFS-LTD) in brain slices from both male and female rats. The metabolic uncoupler fluorocitrate (FC) reduced the probability of transmitter release and depressed synaptic output in a manner that was independent on cannabinoid 1 receptor (CB1R) activation. Fluorocitrate did not affect the LTD induced by the CB1R agonist WIN55,212-2, but enhanced CB1R-dependent HFS-LTD. Reduced neurotransmission and facilitated HFS-LTD were also observed during chemogenetic manipulation using Gi-coupled DREADDs targeting glial fibrillary acidic protein (GFAP)-expressing cells, during the pharmacological inhibition of connexins using carbenoxolone disodium, or during astrocytic glutamate uptake using TFB-TBOA. While pretreatment with the N-methyl-D-aspartate (NMDA) receptor antagonist 2-amino-5-phosphonopentanoic acid (APV) failed to prevent synaptic depression induced by FC, it blocked the facilitation of HFS-LTD. While the lack of tools to disentangle astrocytes from neurons is a major limitation of this study, our data collectively support a role for astrocytes in modulating basal neurotransmission and eCB-mediated synaptic plasticity.
Keywords: LTD; astrocyte; astroglia; endocannabinoid system; synaptic plasticity.
Conflict of interest statement
Authors declare no conflict of interest.
Figures
Similar articles
-
Astrocytes modulate extracellular neurotransmitter levels and excitatory neurotransmission in dorsolateral striatum via dopamine D2 receptor signaling.Neuropsychopharmacology. 2022 Jul;47(8):1493-1502. doi: 10.1038/s41386-021-01232-x. Epub 2021 Nov 22. Neuropsychopharmacology. 2022. PMID: 34811469 Free PMC article.
-
Acute and chronic modulation of striatal endocannabinoid-mediated plasticity by nicotine.Addict Biol. 2019 May;24(3):355-363. doi: 10.1111/adb.12598. Epub 2018 Jan 2. Addict Biol. 2019. PMID: 29292565 Free PMC article.
-
Intermittent ethanol consumption depresses endocannabinoid-signaling in the dorsolateral striatum of rat.Neuropharmacology. 2011 Dec;61(7):1160-5. doi: 10.1016/j.neuropharm.2011.01.014. Epub 2011 Jan 18. Neuropharmacology. 2011. PMID: 21251919
-
Endocannabinoid signaling and long-term synaptic plasticity.Annu Rev Physiol. 2009;71:283-306. doi: 10.1146/annurev.physiol.010908.163149. Annu Rev Physiol. 2009. PMID: 19575681 Free PMC article. Review.
-
Endocannabinoid signaling in synaptic function.Glia. 2023 Jan;71(1):36-43. doi: 10.1002/glia.24256. Epub 2022 Aug 8. Glia. 2023. PMID: 36408881 Free PMC article. Review.
Cited by
-
The Dose-Dependent Effects of Fluorocitrate on the Metabolism and Activity of Astrocytes and Neurons.Brain Sci. 2025 Jan 21;15(2):99. doi: 10.3390/brainsci15020099. Brain Sci. 2025. PMID: 40002432 Free PMC article.
-
Sleep, Glial Function, and the Endocannabinoid System: Implications for Neuroinflammation and Sleep Disorders.Int J Mol Sci. 2024 Mar 9;25(6):3160. doi: 10.3390/ijms25063160. Int J Mol Sci. 2024. PMID: 38542134 Free PMC article. Review.
References
-
- Hansson E., Muyderman H., Leonova J., Allansson L., Sinclair J., Blomstrand F., Thorlin T., Nilsson M., Ronnback L. Astroglia and glutamate in physiology and pathology: Aspects on glutamate transport, glutamate-induced cell swelling and gap-junction communication. Neurochem. Int. 2000;37:317–329. doi: 10.1016/S0197-0186(00)00033-4. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
