

Aerobic glycolysis of bronchial epithelial cells rewires Mycoplasma pneumoniae pneumonia and promotes bacterial elimination
- PMID: 38205952
- PMCID: PMC10863416
- DOI: 10.1128/iai.00248-23
Aerobic glycolysis of bronchial epithelial cells rewires Mycoplasma pneumoniae pneumonia and promotes bacterial elimination
Abstract
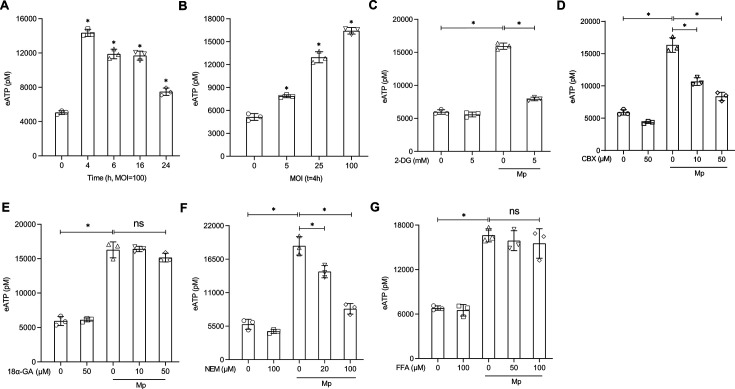
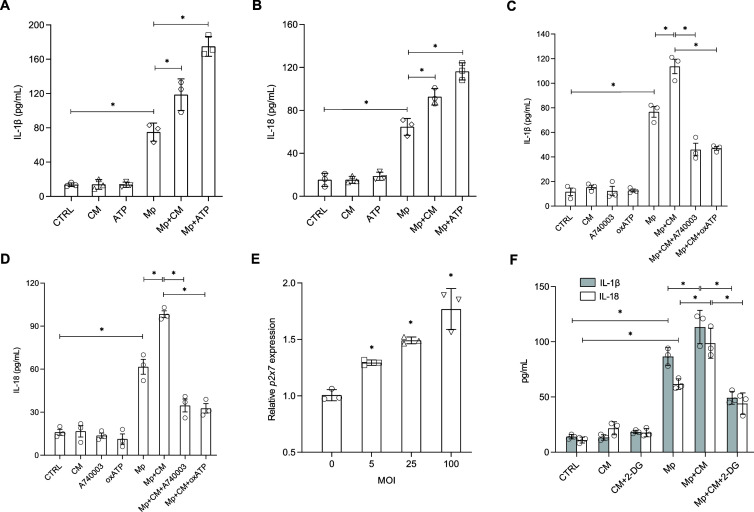
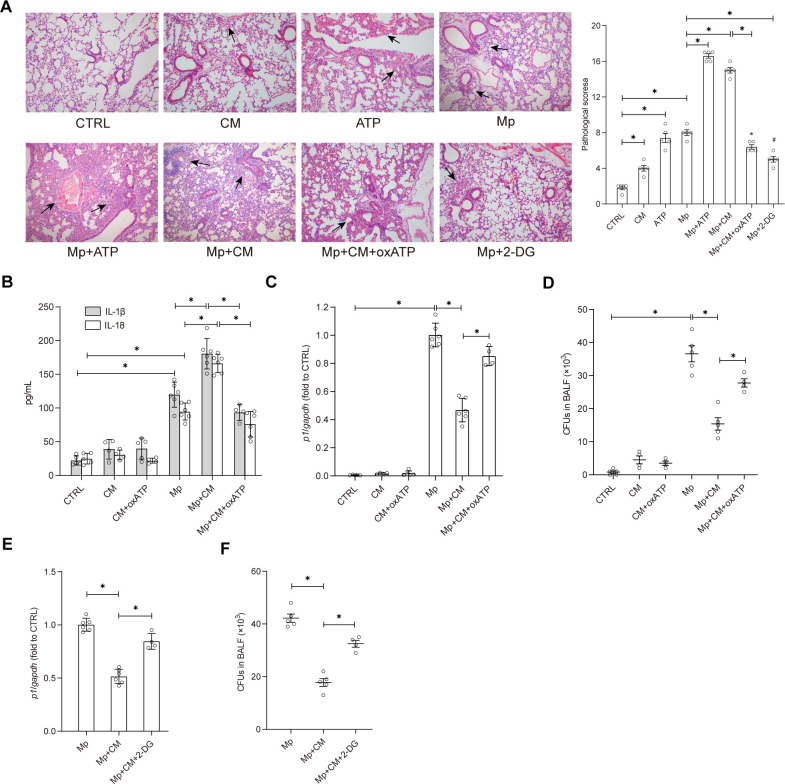
The immune response to Mycoplasma pneumoniae infection plays a key role in clinical symptoms. Previous investigations focused on the pro-inflammatory effects of leukocytes and the pivotal role of epithelial cell metabolic status in finely modulating the inflammatory response have been neglected. Herein, we examined how glycolysis in airway epithelial cells is affected by M. pneumoniae infection in an in vitro model. Additionally, we investigated the contribution of ATP to pulmonary inflammation. Metabolic analysis revealed a marked metabolic shift in bronchial epithelial cells during M. pneumoniae infection, characterized by increased glucose uptake, enhanced aerobic glycolysis, and augmented ATP synthesis. Notably, these metabolic alterations are orchestrated by adaptor proteins, MyD88 and TRAM. The resulting synthesized ATP is released into the extracellular milieu via vesicular exocytosis and pannexin protein channels, leading to a substantial increase in extracellular ATP levels. The conditioned medium supernatant from M. pneumoniae-infected epithelial cells enhances the secretion of both interleukin (IL)-1β and IL-18 by peripheral blood mononuclear cells, partially mediated by the P2X7 purine receptor (P2X7R). In vivo experiments confirm that addition of a conditioned medium exacerbates pulmonary inflammation, which can be attenuated by pre-treatment with a P2X7R inhibitor. Collectively, these findings highlight the significance of airway epithelial aerobic glycolysis in enhancing the pulmonary inflammatory response and aiding pathogen clearance.
Keywords: Mycoplasma pneumoniae; aerobic glycolysis; bronchial epithelial cells; extracellular ATP; peripheral blood mononuclear cells; purine receptor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Targeting the STAT3/ACLY axis attenuates pulmonary inflammation but delays Mycoplasma pneumoniae clearance via citrate metabolism.Med Microbiol Immunol. 2025 May 28;214(1):26. doi: 10.1007/s00430-025-00836-0. Med Microbiol Immunol. 2025. PMID: 40434443
-
NLRP3 Is a Critical Regulator of Inflammation and Innate Immune Cell Response during Mycoplasma pneumoniae Infection.Infect Immun. 2017 Dec 19;86(1):e00548-17. doi: 10.1128/IAI.00548-17. Print 2018 Jan. Infect Immun. 2017. PMID: 29061706 Free PMC article.
-
Release of cytokines by human nasal epithelial cells and peripheral blood mononuclear cells infected with Mycoplasma pneumoniae.Exp Biol Med (Maywood). 2002 May;227(5):330-5. doi: 10.1177/153537020222700505. Exp Biol Med (Maywood). 2002. PMID: 11976403
-
Interaction between alveolar macrophages and epithelial cells during Mycoplasma pneumoniae infection.Front Cell Infect Microbiol. 2023 Apr 11;13:1052020. doi: 10.3389/fcimb.2023.1052020. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37113130 Free PMC article. Review.
-
Pseudomembranous necrotizing laryngotracheobronchitis due to Mycoplasma pneumoniae: a case report and literature review.BMC Infect Dis. 2022 Feb 23;22(1):183. doi: 10.1186/s12879-022-07160-5. BMC Infect Dis. 2022. PMID: 35197010 Free PMC article. Review.
Cited by
-
Risk factors for refractory Mycoplasma pneumoniae in Chinese children: a meta-analysis.Front Pediatr. 2025 Jun 27;13:1512689. doi: 10.3389/fped.2025.1512689. eCollection 2025. Front Pediatr. 2025. PMID: 40656195 Free PMC article.
-
Unraveling the role of distinct cytoskeletal motility structures in Mycoplasma pneumoniae relatives.BMC Microbiol. 2025 Aug 29;25(1):562. doi: 10.1186/s12866-025-04320-w. BMC Microbiol. 2025. PMID: 40883708 Review.
-
MreB: unraveling the molecular mechanisms of bacterial shape, division, and environmental adaptation.Cell Commun Signal. 2025 Aug 22;23(1):377. doi: 10.1186/s12964-025-02373-y. Cell Commun Signal. 2025. PMID: 40847400 Free PMC article. Review.
-
Targeting the STAT3/ACLY axis attenuates pulmonary inflammation but delays Mycoplasma pneumoniae clearance via citrate metabolism.Med Microbiol Immunol. 2025 May 28;214(1):26. doi: 10.1007/s00430-025-00836-0. Med Microbiol Immunol. 2025. PMID: 40434443
-
Revealing the impact of Pseudomonas aeruginosa quorum sensing molecule 2'-aminoacetophenone on the human bronchial-airway epithelium and pulmonary endothelium using a human airway-on-a-chip.Front Immunol. 2025 Jul 15;16:1592597. doi: 10.3389/fimmu.2025.1592597. eCollection 2025. Front Immunol. 2025. PMID: 40735328 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- 81902077, 82101869/MOST | National Natural Science Foundation of China (NSFC)
- 2022JJ30488, 2022JJ40406/Natural Science Foundation of Hunan Province, China
- 2021SK51906/Hunan Province Science and Technology Innovation Plant Project
- 20201915/Scientific Research Project of Hunan Provincial Health Commitee
- 20B498/Research Foundation of Education Bureau of Hunan Province
- CX20200958, CX202211004/Graduate student scientific research innovation Projects in Hunan Province
- ptkwws202314/Technology Innovation Project Plan of Health System of Shanghai Putuo District
- 2022rmky02/Research Project Plan of the People's Hospital of Putuo District, Shanghai
- 202250045307/Hengyang Science and Technology Planning Project
- NO.12/the Emergency special project of epidemic prevention and control of COVID-19 pneumonia in the Unoversity of South China
LinkOut - more resources
Full Text Sources
Miscellaneous
