

Increased privatization of a public resource leads to spread of cooperation in a microbial population
- PMID: 38206031
- PMCID: PMC10846273
- DOI: 10.1128/spectrum.02358-23
Increased privatization of a public resource leads to spread of cooperation in a microbial population
Abstract
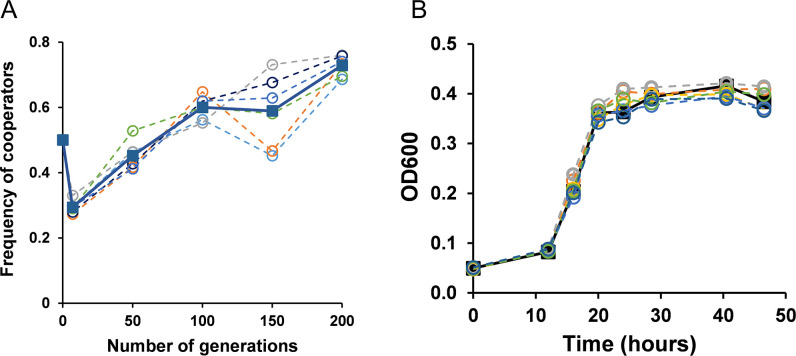
The phenomenon of cooperation is prevalent at all levels of life. In one such manifestation of cooperation in microbial communities, some cells produce costly extracellular resources that are freely available to others. These resources are referred to as public goods. Saccharomyces cerevisiae secretes invertase (public good) in the periplasm to hydrolyze sucrose into glucose and fructose, which are then imported by the cells. After hydrolysis of sucrose, a cooperator retains only 1% of the monosaccharides, while 99% of the monosaccharides diffuse into the environment and can be utilized by any cell. The non-producers of invertase (cheaters) exploit the invertase-producing cells (cooperators) by utilizing the monosaccharides and not paying the metabolic cost of producing the invertase. In this work, we investigate the evolutionary dynamics of this cheater-cooperator system. In a co-culture, if cheaters are selected for their higher fitness, the population will collapse. On the other hand, for cooperators to survive in the population, a strategy to increase fitness would likely be required. To understand the adaptation of cooperators in sucrose, we performed a coevolution experiment in sucrose. Our results show that cooperators increase in fitness as the experiment progresses. This phenomenon was not observed in environments which involved a non-public good system. Genome sequencing reveals duplication of several HXT transporters in the evolved cooperators. Based on these results, we hypothesize that increased privatization of the monosaccharides is the most likely explanation of spread of cooperators in the population.IMPORTANCEHow is cooperation, as a trait, maintained in a population? In order to answer this question, we perform a coevolution experiment between two strains of yeast-one which produces a public good to release glucose and fructose in the media, thus generating a public resource, and the other which does not produce public resource and merely benefits from the presence of the cooperator strain. What is the outcome of this coevolution experiment? We demonstrate that after ~200 generations of coevolution, cooperators increase in frequency in the co-culture. Remarkably, in all parallel lines of our experiment, this is obtained via duplication of regions which likely allow greater privatization of glucose and fructose. Thus, increased privatization, which is intuitively thought to be a strategy against cooperation, enables spread of cooperation.
Keywords: CNV; SNV/indel; cheating; cooperation; microbe; public good system; sucrose; yeast.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
Snowdrift game dynamics and facultative cheating in yeast.Nature. 2009 May 14;459(7244):253-6. doi: 10.1038/nature07921. Epub 2009 Apr 6. Nature. 2009. PMID: 19349960 Free PMC article.
-
Optogenetic spatial patterning of cooperation in yeast populations.Nat Commun. 2024 Jan 2;15(1):75. doi: 10.1038/s41467-023-44379-5. Nat Commun. 2024. PMID: 38168087 Free PMC article.
-
Adaptation to a new environment allows cooperators to purge cheaters stochastically.Proc Natl Acad Sci U S A. 2012 Nov 20;109(47):19079-86. doi: 10.1073/pnas.1210190109. Epub 2012 Oct 22. Proc Natl Acad Sci U S A. 2012. PMID: 23091010 Free PMC article.
-
Cheating and punishment in cooperative animal societies.Philos Trans R Soc Lond B Biol Sci. 2016 Feb 5;371(1687):20150090. doi: 10.1098/rstb.2015.0090. Philos Trans R Soc Lond B Biol Sci. 2016. PMID: 26729930 Free PMC article. Review.
-
Evolution of Sucrose Metabolism: The Dichotomy of Invertases and Beyond.Trends Plant Sci. 2018 Feb;23(2):163-177. doi: 10.1016/j.tplants.2017.11.001. Epub 2017 Nov 25. Trends Plant Sci. 2018. PMID: 29183781 Review.
Cited by
-
Empirical evidence of resource dependent evolution of payoff matrices in Saccharomyces cerevisiae populations.J Evol Biol. 2025 Jan 3;38(1):122-128. doi: 10.1093/jeb/voae128. J Evol Biol. 2025. PMID: 39387146
-
Resource presentation dictates genetic and phenotypic adaptation in yeast.BMC Ecol Evol. 2025 Apr 15;25(1):33. doi: 10.1186/s12862-025-02361-3. BMC Ecol Evol. 2025. PMID: 40234742 Free PMC article.
-
Microbial cross-feeding stabilized by segregation of a dependent mutant from its independent ancestor.ISME J. 2025 Jan 2;19(1):wraf131. doi: 10.1093/ismejo/wraf131. ISME J. 2025. PMID: 40568968 Free PMC article.
References
-
- West SA, Diggle SP, Buckling A, Gardner A, Griffin AS. 2007. The social lives of microbes. Annu Rev Ecol Evol Syst 38:53–77. doi:10.1146/annurev.ecolsys.38.091206.095740 - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
