

Post-pandemic memory T cell response to SARS-CoV-2 is durable, broadly targeted, and cross-reactive to the hypermutated BA.2.86 variant
- PMID: 38211583
- PMCID: PMC10901529
- DOI: 10.1016/j.chom.2023.12.003
Post-pandemic memory T cell response to SARS-CoV-2 is durable, broadly targeted, and cross-reactive to the hypermutated BA.2.86 variant
Abstract
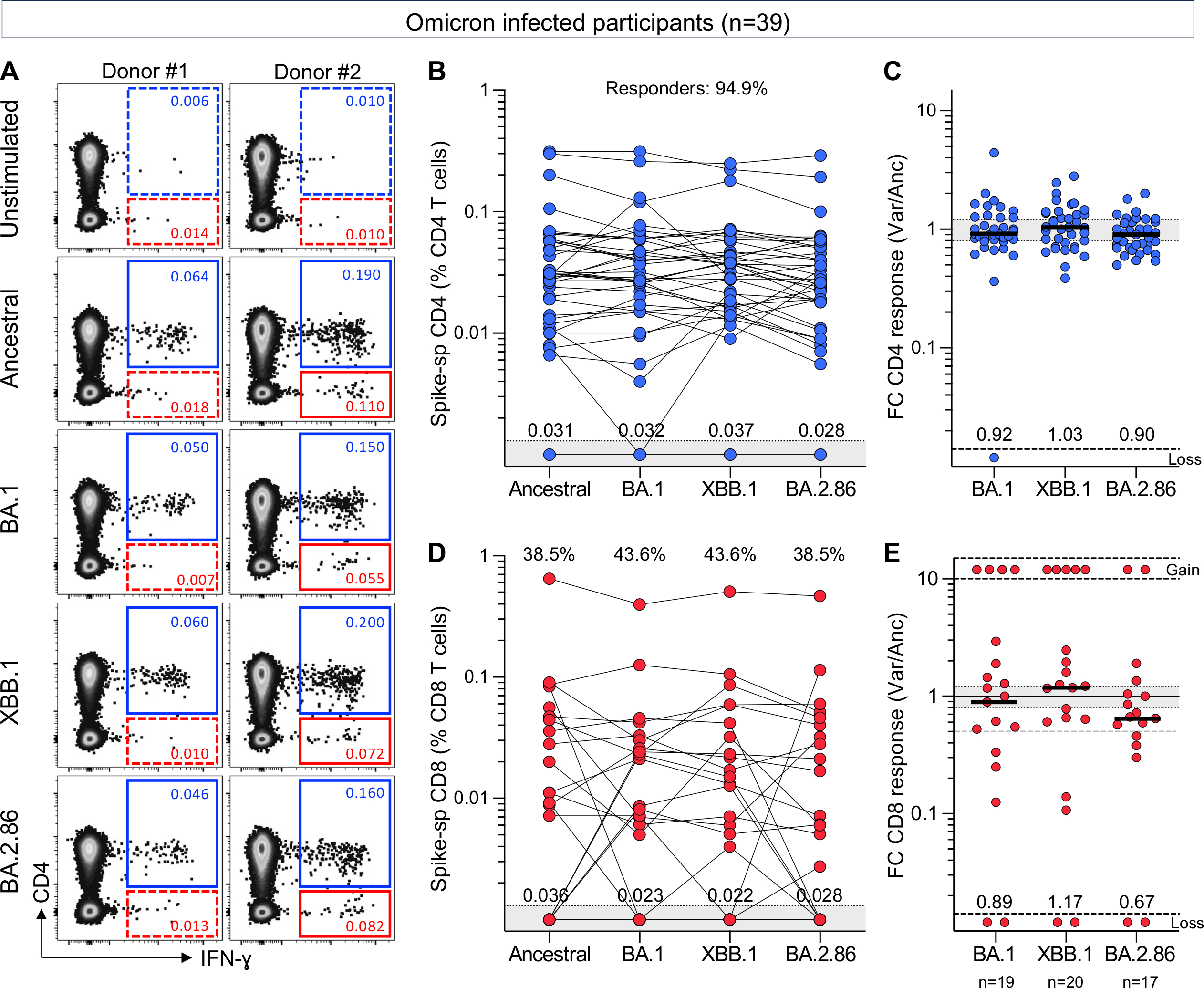
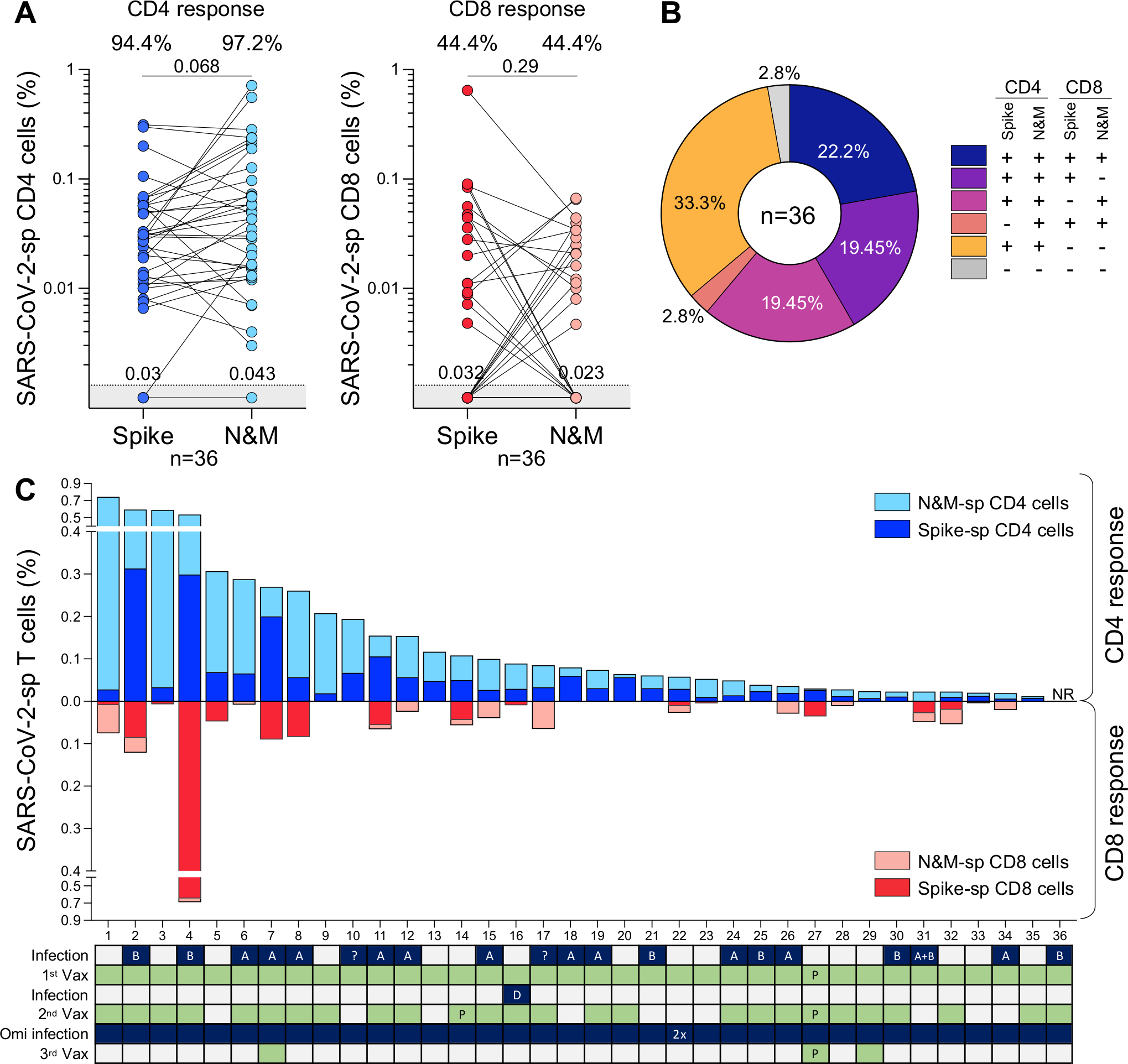
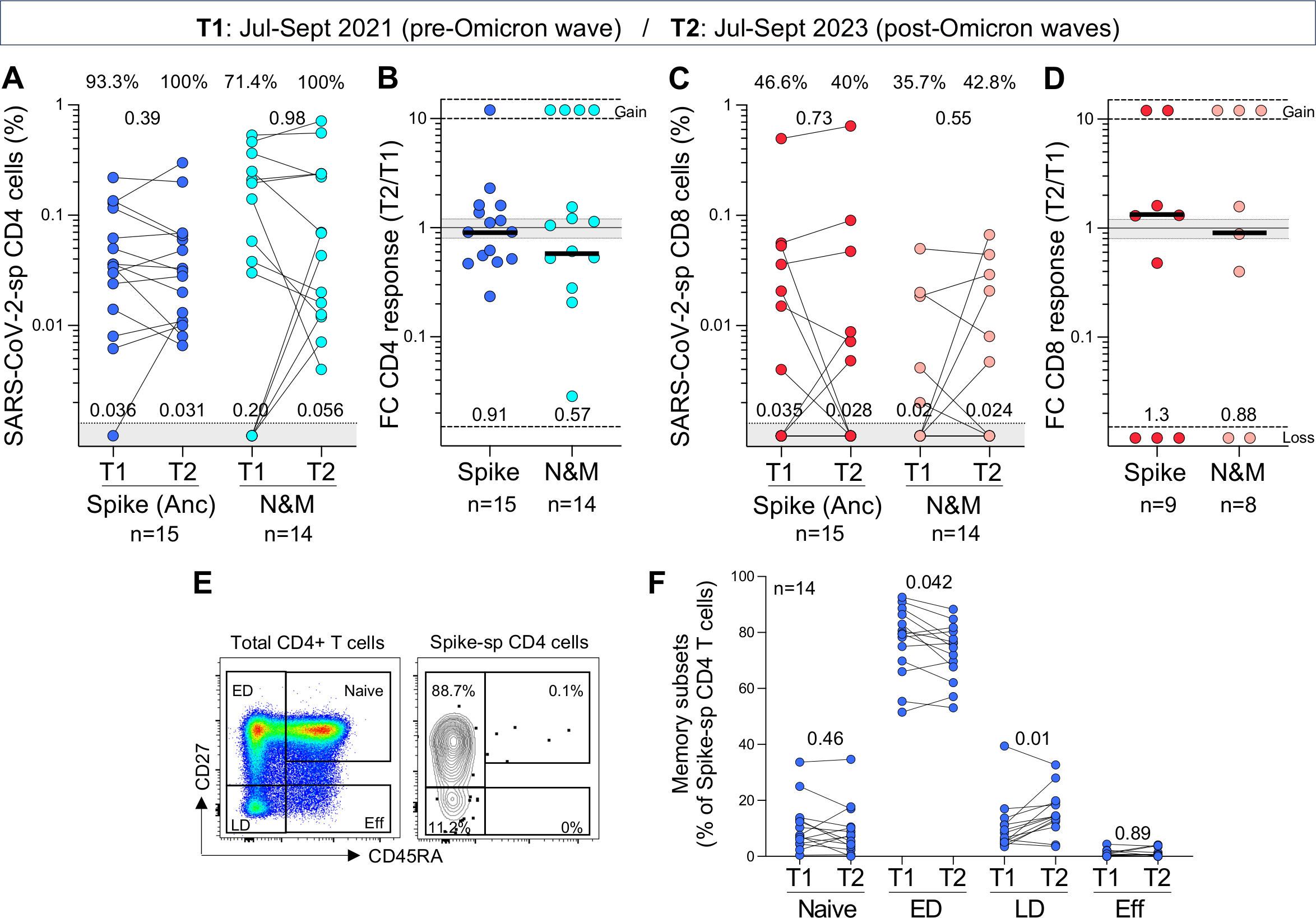
Ongoing severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) evolution has given rise to recombinant Omicron lineages that dominate globally (XBB.1), as well as the emergence of hypermutated variants (BA.2.86). In this context, durable and cross-reactive T cell immune memory is critical for continued protection against severe COVID-19. We examined T cell responses to SARS-CoV-2 approximately 1.5 years since Omicron first emerged. We describe sustained CD4+ and CD8+ spike-specific T cell memory responses in healthcare workers in South Africa (n = 39) who were vaccinated and experienced at least one SARS-CoV-2 infection. Spike-specific T cells are highly cross-reactive with all Omicron variants tested, including BA.2.86. Abundant nucleocapsid and membrane-specific T cells are detectable in most participants. The bulk of SARS-CoV-2-specific T cell responses have an early-differentiated phenotype, explaining their persistent nature. Overall, hybrid immunity leads to the accumulation of spike and non-spike T cells evident 3.5 years after the start of the pandemic, with preserved recognition of highly mutated SARS-CoV-2 variants.
Keywords: BA.2.86 sub-lineage; SARS-CoV-2; T cell response; T cell sustainability; cross-reactivity.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.S. is a consultant for AstraZeneca Pharmaceuticals, Calyptus Pharmaceuticals Inc, Darwin Health, EmerVax, EUROIMMUN, F. Hoffman-La Roche Ltd, Fortress Biotech, Gilead Sciences, Granite bio., Gritstone Oncology, Guggenheim Securities, Moderna, Pfizer, RiverVest Venture Partners, and Turnstone Biologics. A.G. is a consultant for Pfizer. L.J.I. has filed for patent protection for various aspects of T cell epitope and vaccine design work.
Figures
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
