

A comparative evaluation of the strengths and potential caveats of the microglial inducible CreER mouse models
- PMID: 38217856
- PMCID: PMC10874587
- DOI: 10.1016/j.celrep.2023.113660
A comparative evaluation of the strengths and potential caveats of the microglial inducible CreER mouse models
Abstract
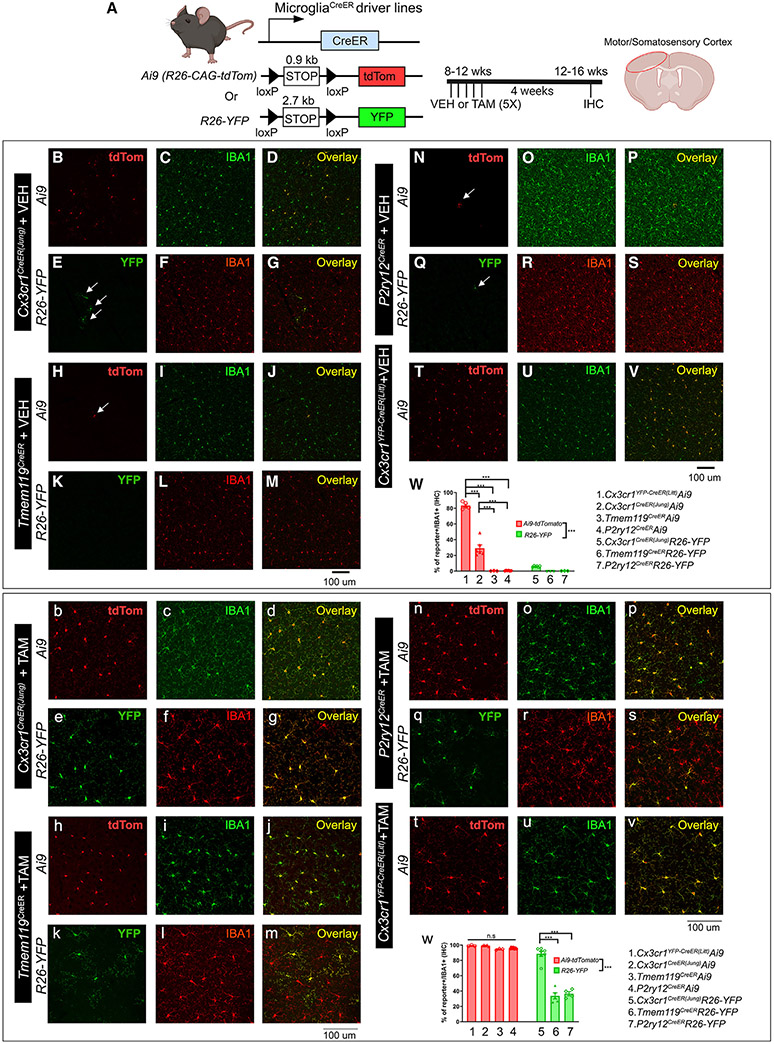
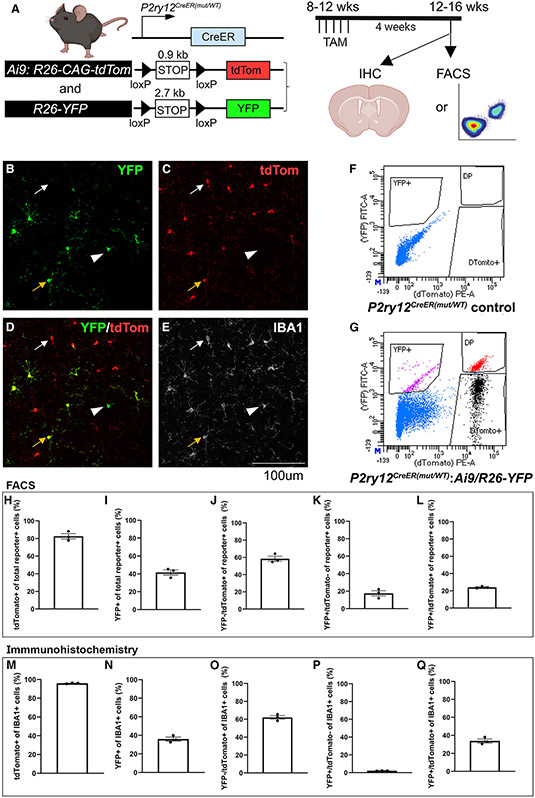
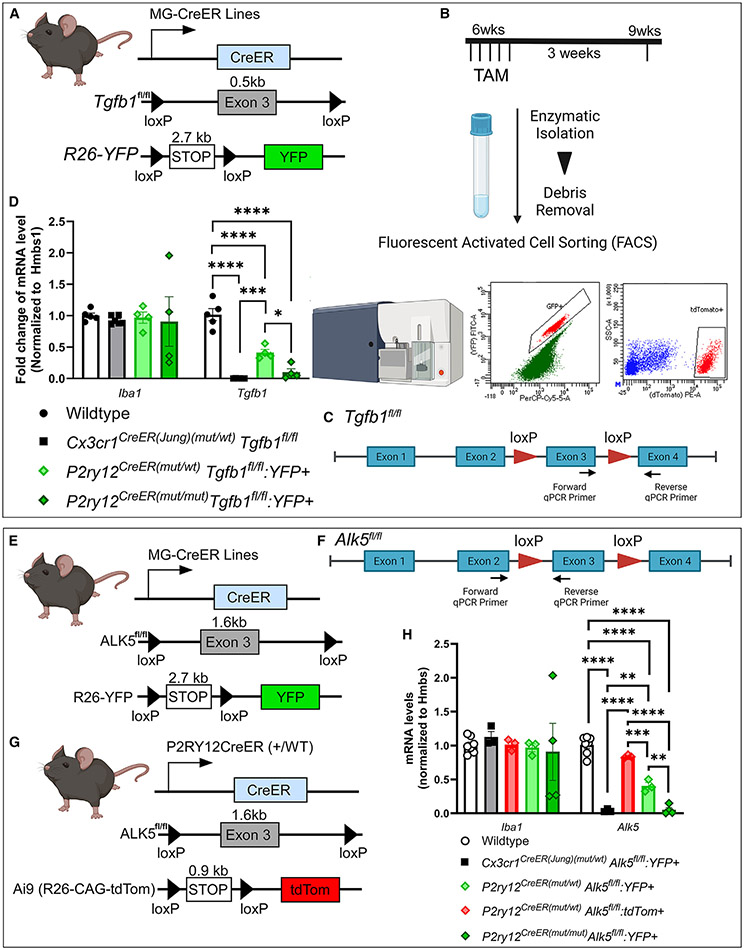
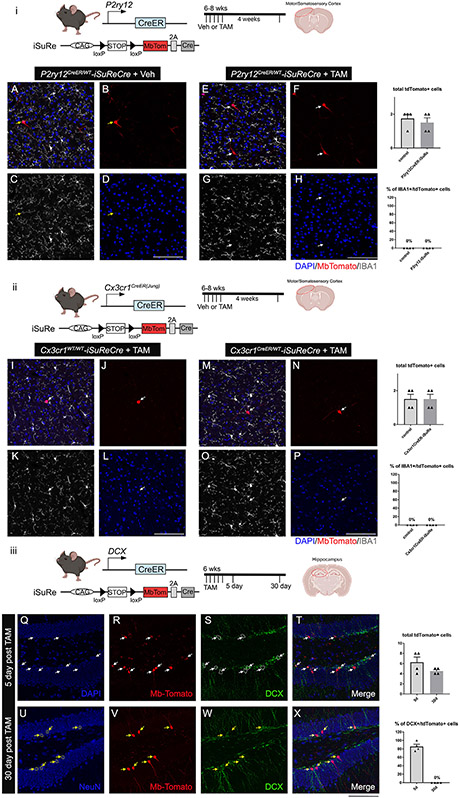
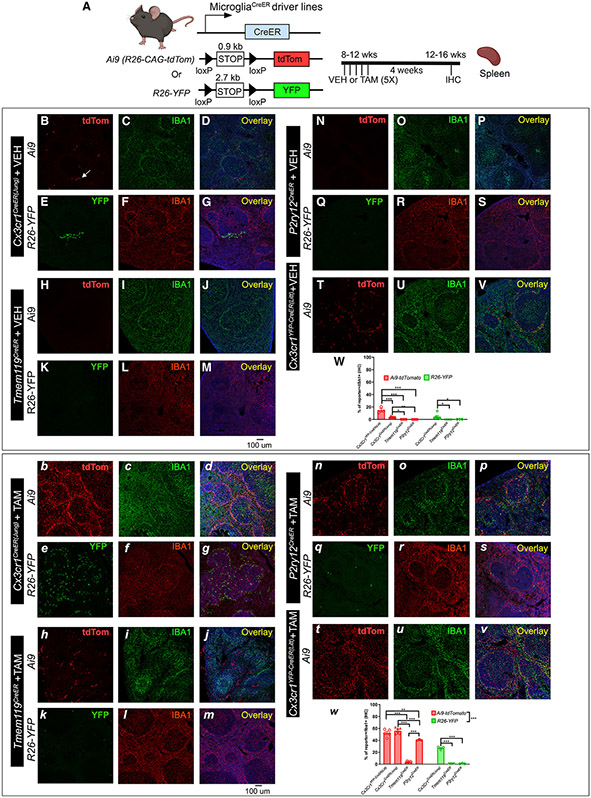
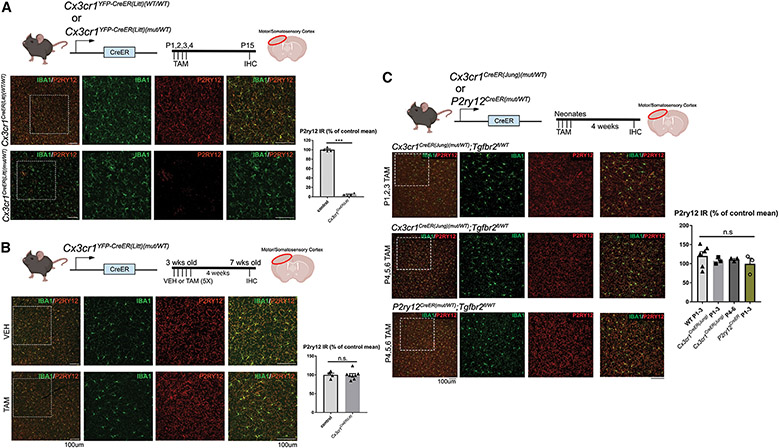
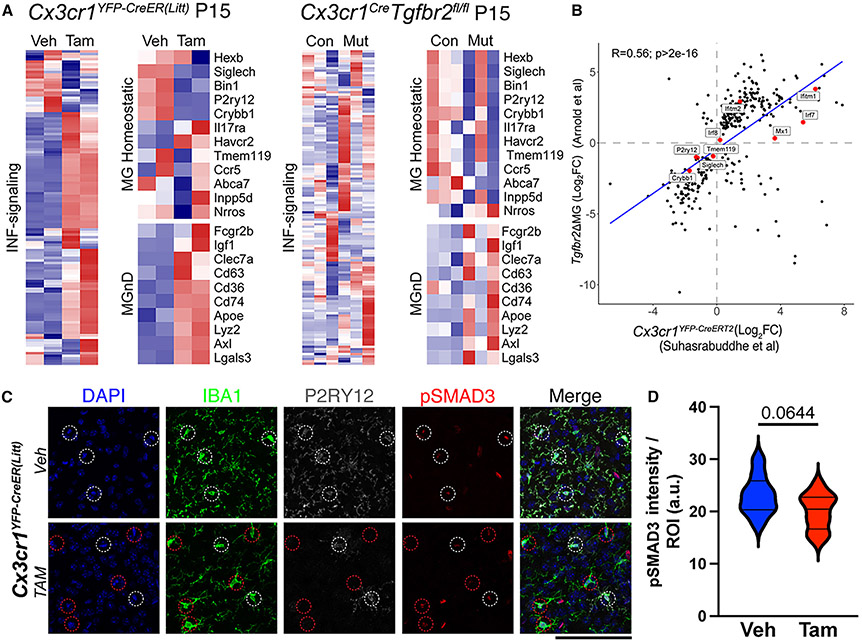
The recent proliferation of new Cre and CreER recombinase lines provides researchers with a diverse toolkit to study microglial gene function. To determine how best to apply these lines in studies of microglial gene function, a thorough and detailed comparison of their properties is needed. Here, we examined four different microglial CreER lines (Cx3cr1YFP-CreER(Litt), Cx3cr1CreER(Jung), P2ry12CreER, and Tmem119CreER), focusing on (1) recombination specificity, (2) leakiness (the degree of tamoxifen-independent recombination in microglia and other cells), (3) the efficiency of tamoxifen-induced recombination, (4) extraneural recombination (the degree of recombination in cells outside of the CNS, particularly myelo/monocyte lineages), and (5) off-target effects in the context of neonatal brain development. We identify important caveats and strengths for these lines, which will provide broad significance for researchers interested in performing conditional gene deletion in microglia. We also provide data emphasizing the potential of these lines for injury models that result in the recruitment of splenic immune cells.
Keywords: CP: Neuroscience; CX3CR1; Cre recombinase; P2RY12; inducible gene recombination; microglia; tmem119.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Update of
-
Finding the right tool: a comprehensive evaluation of microglial inducible cre mouse models.bioRxiv [Preprint]. 2023 Apr 17:2023.04.17.536878. doi: 10.1101/2023.04.17.536878. bioRxiv. 2023. Update in: Cell Rep. 2024 Jan 23;43(1):113660. doi: 10.1016/j.celrep.2023.113660. PMID: 37131606 Free PMC article. Updated. Preprint.
References
-
- Nimmerjahn A, Kirchhoff F, and Helmchen F (2005). Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 308, 1314–1318. - PubMed
-
- Paolicelli RC, Bolasco G, Pagani F, Maggi L, Scianni M, Panzanelli P, Giustetto M, Ferreira TA, Guiducci E, Dumas L, et al. (2011). Synaptic pruning by microglia is necessary for normal brain development. Science 333, 1456–1458. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
