

Versatile CYP98A enzymes catalyse meta-hydroxylation reveals diversity of salvianolic acids biosynthesis
- PMID: 38226779
- PMCID: PMC11123398
- DOI: 10.1111/pbi.14284
Versatile CYP98A enzymes catalyse meta-hydroxylation reveals diversity of salvianolic acids biosynthesis
Abstract
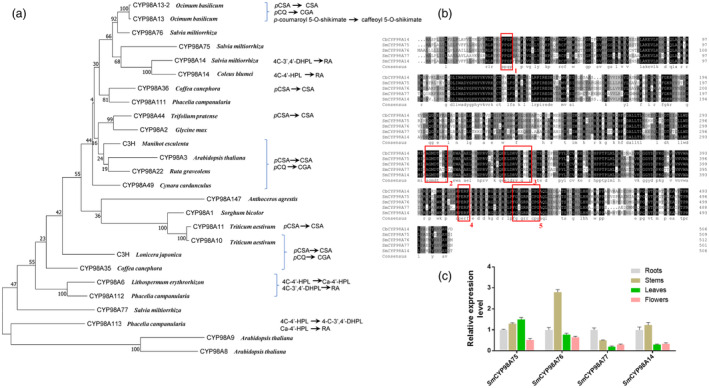
Salvianolic acids (SA), such as rosmarinic acid (RA), danshensu (DSS), and their derivative salvianolic acid B (SAB), etc. widely existed in Lamiaceae and Boraginaceae families, are of interest due to medicinal properties in the pharmaceutical industries. Hundreds of studies in past decades described that 4-coumaroyl-CoA and 4-hydroxyphenyllactic acid (4-HPL) are common substrates to biosynthesize SA with participation of rosmarinic acid synthase (RAS) and cytochrome P450 98A (CYP98A) subfamily enzymes in different plants. However, in our recent study, several acyl donors and acceptors included DSS as well as their ester-forming products all were determined in SA-rich plants, which indicated that previous recognition to SA biosynthesis is insufficient. Here, we used Salvia miltiorrhiza, a representative important medicinal plant rich in SA, to elucidate the diversity of SA biosynthesis. Various acyl donors as well as acceptors are catalysed by SmRAS to form precursors of RA and two SmCYP98A family members, SmCYP98A14 and SmCYP98A75, are responsible for different positions' meta-hydroxylation of these precursors. SmCYP98A75 preferentially catalyses C-3' hydroxylation, and SmCYP98A14 preferentially catalyses C-3 hydroxylation in RA generation. In addition, relative to C-3' hydroxylation of the acyl acceptor moiety in RA biosynthesis, SmCYP98A75 has been verified as the first enzyme that participates in DSS formation. Furthermore, SmCYP98A enzymes knockout resulted in the decrease and overexpression leaded to dramatic increase of SA accumlation. Our study provides new insights into SA biosynthesis diversity in SA-abundant species and versatility of CYP98A enzymes catalytic preference in meta-hydroxylation reactions. Moreover, CYP98A enzymes are ideal metabolic engineering targets to elevate SA content.
Keywords: CYP98A; biosynthesis; danshensu; rosmarinic acid; salvianolic acid.
© 2024 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures





Similar articles
-
The chromosome-scale assembly of the Salvia plebeia genome provides insight into the biosynthesis and regulation of rosmarinic acid.Plant Biotechnol J. 2025 May;23(5):1507-1520. doi: 10.1111/pbi.14601. Epub 2025 Feb 13. Plant Biotechnol J. 2025. PMID: 39945326 Free PMC article.
-
Gibberellin-Induced Transcription Factor SmMYB71 Negatively Regulates Salvianolic Acid Biosynthesis in Salvia miltiorrhiza.Molecules. 2024 Dec 13;29(24):5892. doi: 10.3390/molecules29245892. Molecules. 2024. PMID: 39769982 Free PMC article.
-
Analysis of rosmarinic acid synthase (RAS) gene family and functional study of SmRAS1/2/4 in Salvia miltiorrhiza.Gene. 2025 Jan 20;934:149057. doi: 10.1016/j.gene.2024.149057. Epub 2024 Oct 26. Gene. 2025. PMID: 39490649
-
Evolution of rosmarinic acid biosynthesis.Phytochemistry. 2009 Oct-Nov;70(15-16):1663-79. doi: 10.1016/j.phytochem.2009.05.010. Epub 2009 Jun 25. Phytochemistry. 2009. PMID: 19560175 Review.
-
Molecular Pharmacology of Rosmarinic and Salvianolic Acids: Potential Seeds for Alzheimer's and Vascular Dementia Drugs.Int J Mol Sci. 2018 Feb 3;19(2):458. doi: 10.3390/ijms19020458. Int J Mol Sci. 2018. PMID: 29401682 Free PMC article. Review.
Cited by
-
UHPLC-QTOF-MS-Based Targeted Metabolomics Provides Novel Insights into the Accumulative Mechanism of Soil Types on the Bioactive Components of Salvia miltiorrhiza.Molecules. 2024 Aug 25;29(17):4016. doi: 10.3390/molecules29174016. Molecules. 2024. PMID: 39274864 Free PMC article.
-
The chromosome-scale assembly of the Salvia plebeia genome provides insight into the biosynthesis and regulation of rosmarinic acid.Plant Biotechnol J. 2025 May;23(5):1507-1520. doi: 10.1111/pbi.14601. Epub 2025 Feb 13. Plant Biotechnol J. 2025. PMID: 39945326 Free PMC article.
-
Ecotype-specific phenolic acid accumulation and root softness in Salvia miltiorrhiza are driven by environmental and genetic factors.Plant Biotechnol J. 2025 Jun;23(6):2224-2241. doi: 10.1111/pbi.70048. Epub 2025 Mar 19. Plant Biotechnol J. 2025. PMID: 40107323 Free PMC article.
-
Isolation of Endophytic Fungi and Effects on Secondary Metabolites in Hairy Roots of Salvia miltiorrhiza.J Microbiol Biotechnol. 2025 Apr 9;35:e2411051. doi: 10.4014/jmb.2411.11051. J Microbiol Biotechnol. 2025. PMID: 40223278 Free PMC article.
-
Analysis of the chemical constituents and their metabolites in Orthosiphon stamineus Benth. via UHPLC-Q exactive orbitrap-HRMS and AFADESI-MSI techniques.PLoS One. 2024 Jun 25;19(6):e0304852. doi: 10.1371/journal.pone.0304852. eCollection 2024. PLoS One. 2024. PMID: 38917120 Free PMC article.
References
-
- Di, P. , Zhang, L. , Chen, J. , Tan, H. , Xiao, Y. , Dong, X. , Zhou, X. et al. (2013) 13C tracer reveals phenolic acids biosynthesis in hairy root cultures of Salvia miltiorrhiza . ACS Chem. Biol. 8, 1537–1548. - PubMed
-
- Eberle, D. , Ullmann, P. , Werck‐Reichhart, D. and Petersen, M. (2009) cDNA cloning and functional characterisation of CYP98A14 and NADPH:cytochrome P450 reductase from Coleus blumei involved in rosmarinic acid biosynthesis. Plant Mol. Biol. 69, 239–253. - PubMed
-
- Gang, D.R. , Beuerle, T. , Ullmann, P. , Werck‐Reichhart, D. and Pichersky, E. (2002) Differential production of meta hydroxylated phenylpropanoids in sweet basil peltate glandular trichomes and leaves is controlled by the activities of specific acyltransferases and hydroxylases. Plant Physiol. 130, 1536–1544. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
