

The effector program of human CD8 T cells supports tissue remodeling
- PMID: 38226976
- PMCID: PMC10791561
- DOI: 10.1084/jem.20230488
The effector program of human CD8 T cells supports tissue remodeling
Abstract
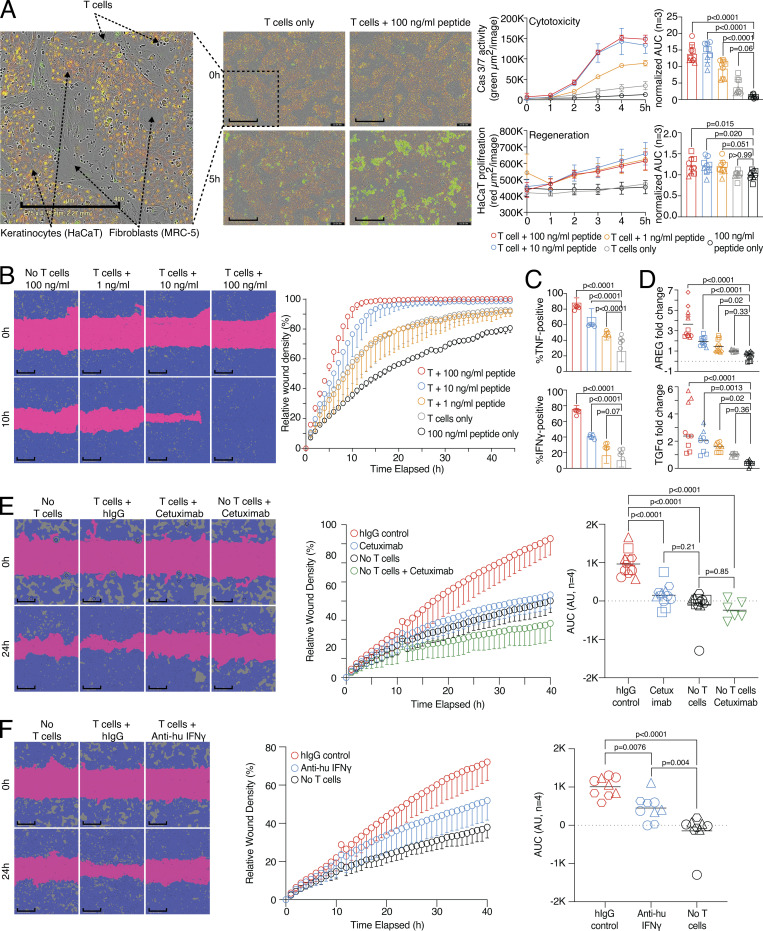
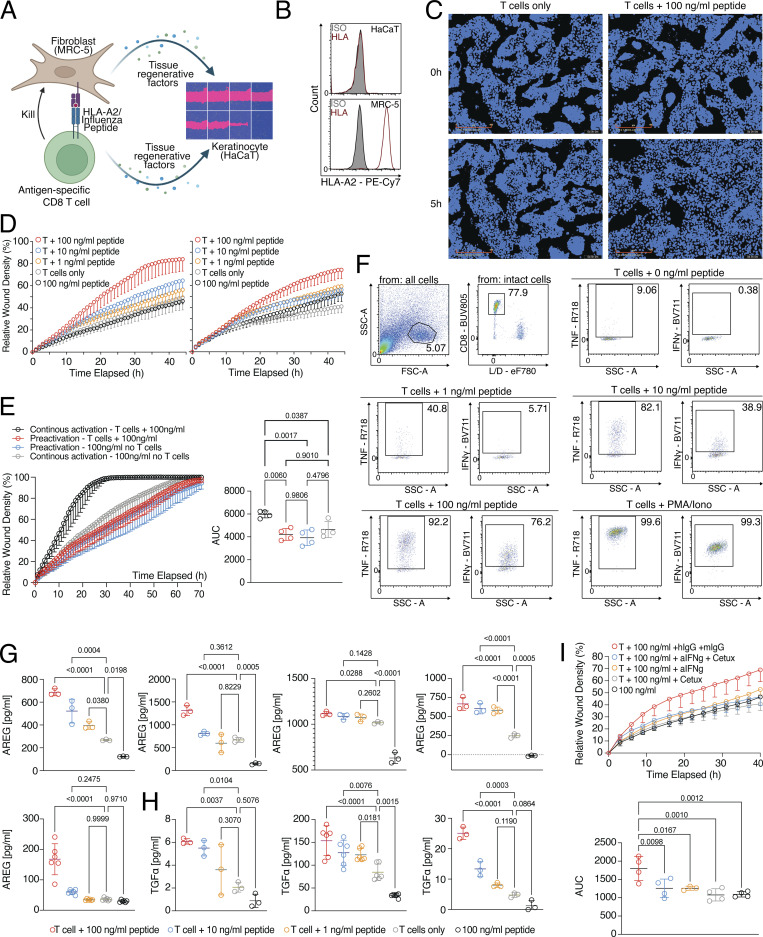
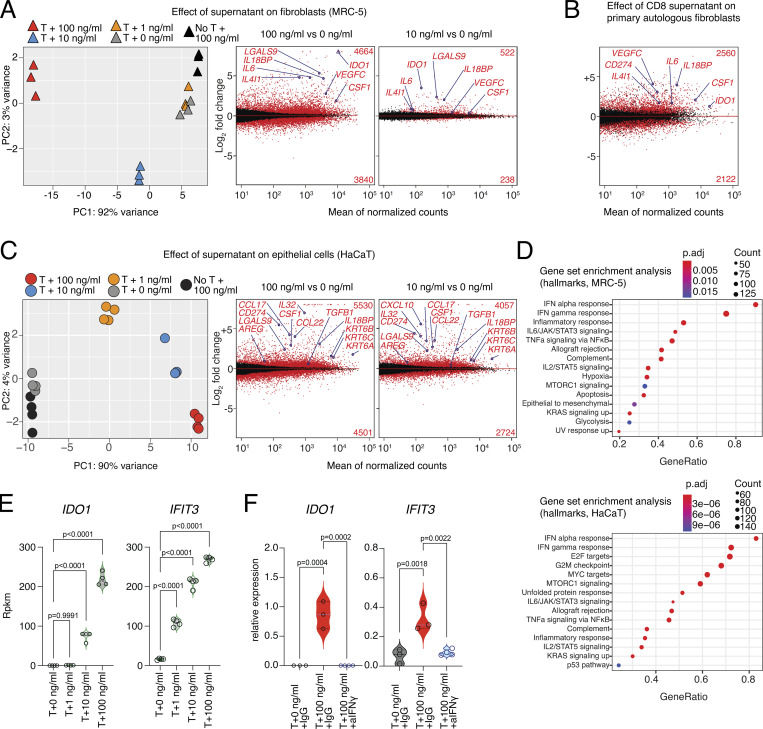
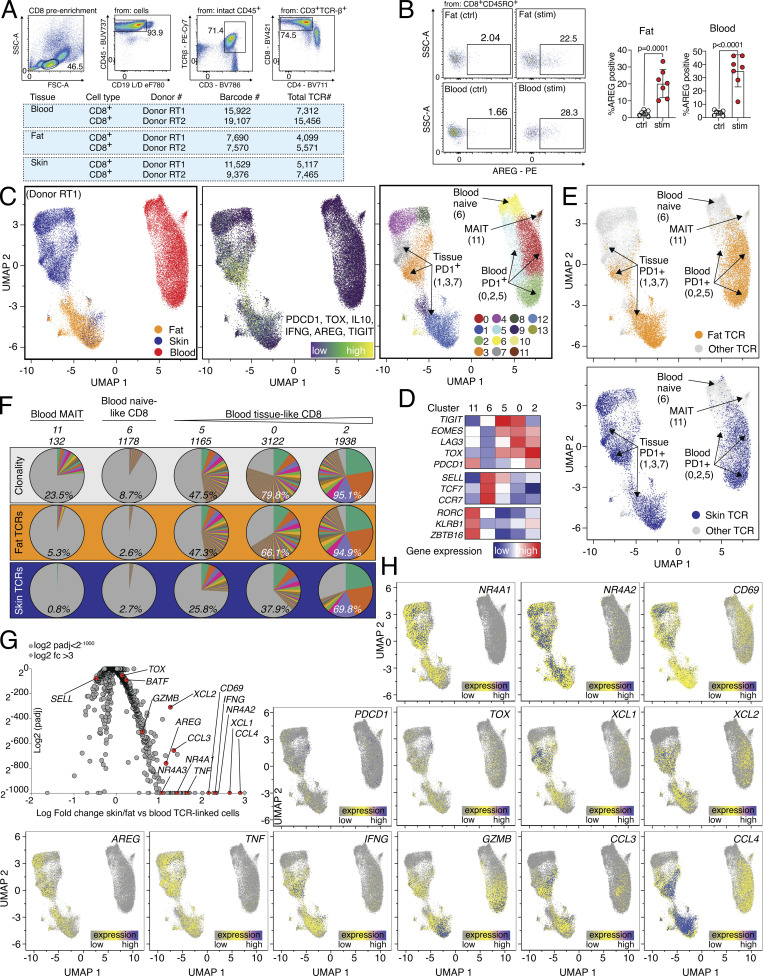
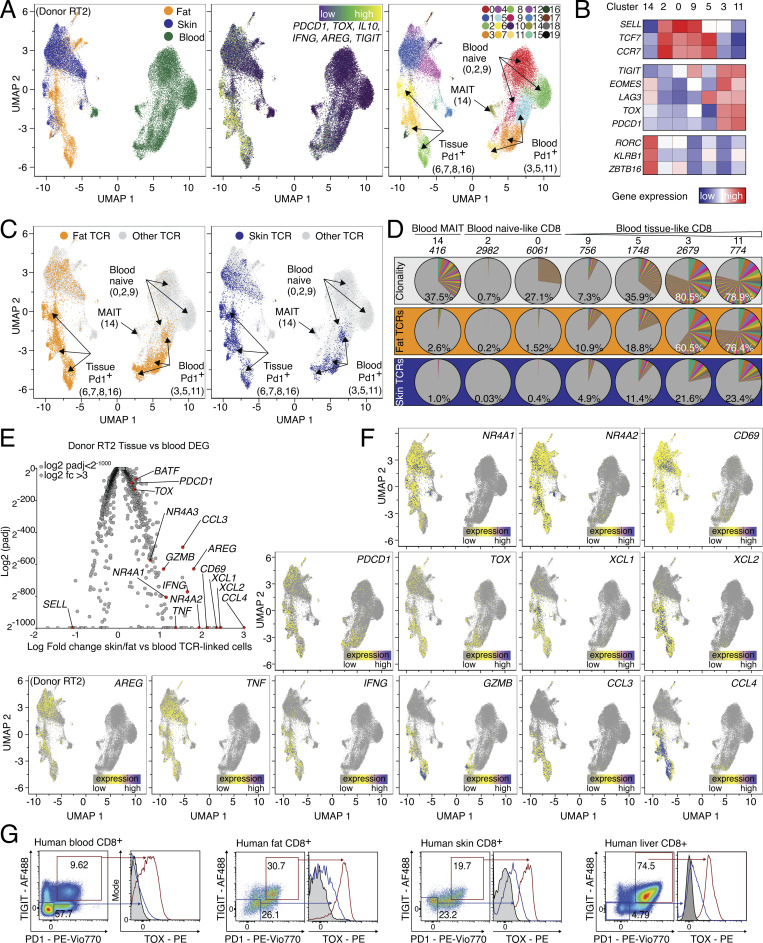
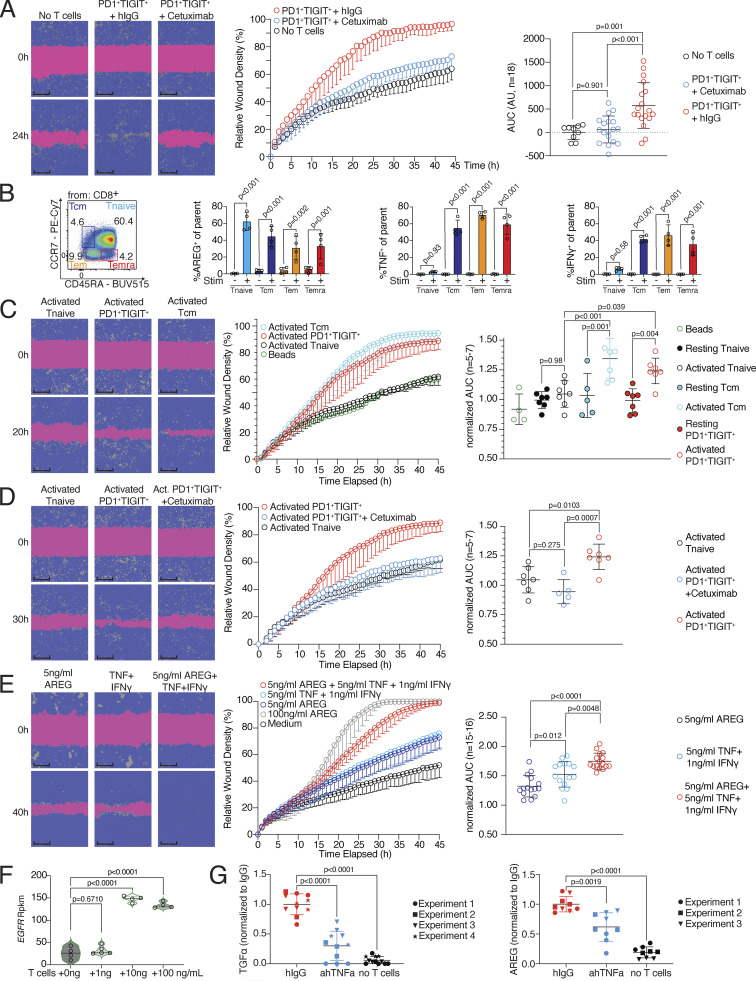
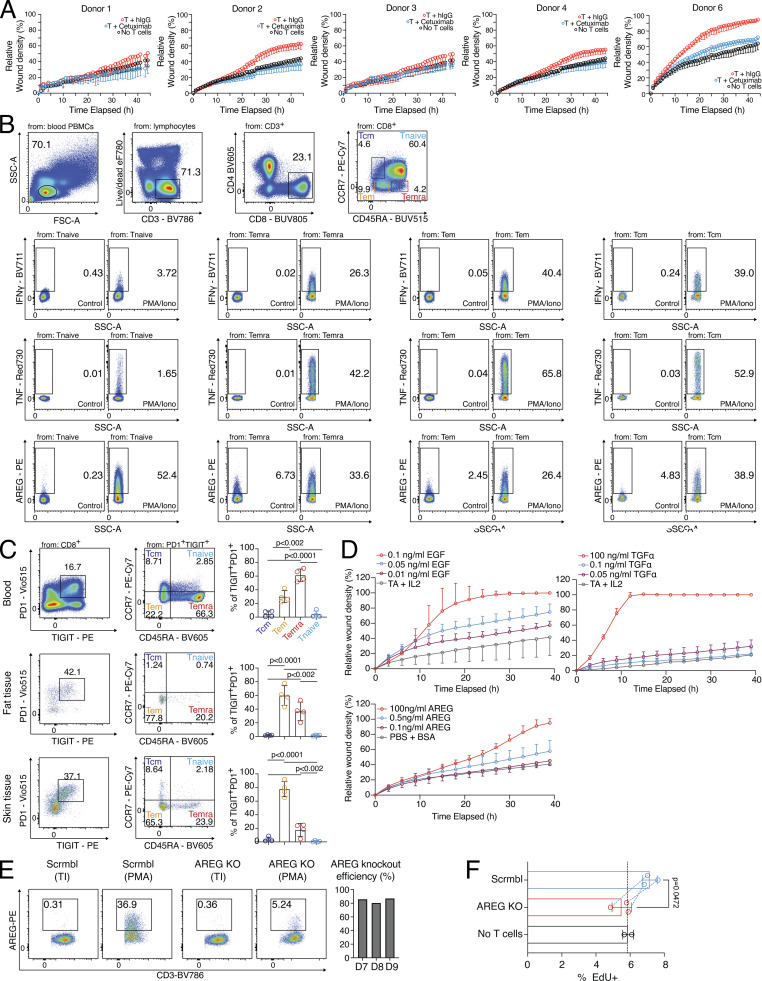
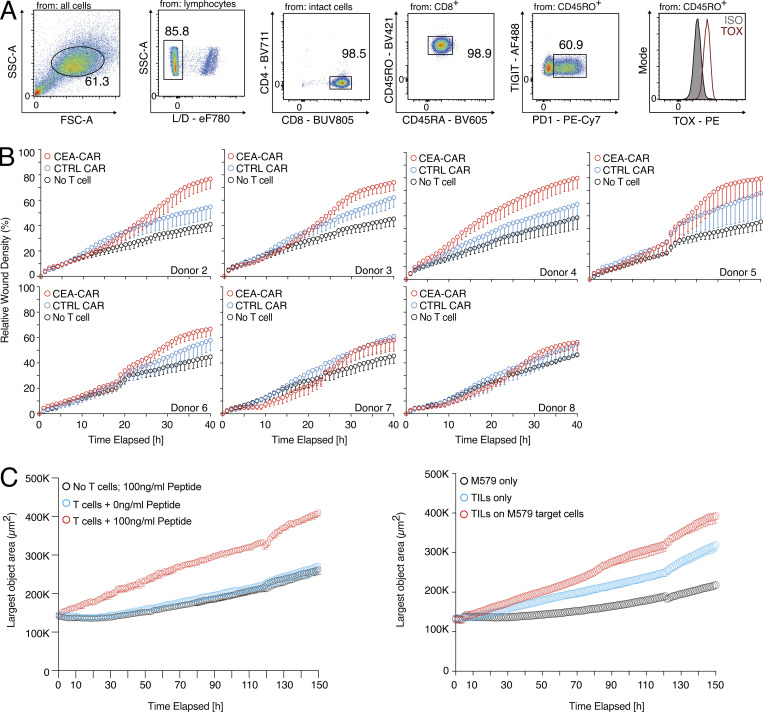
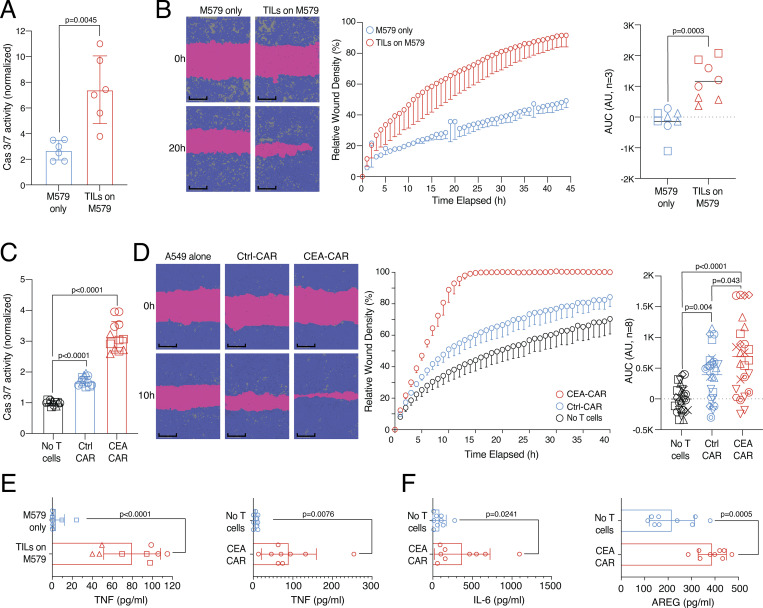
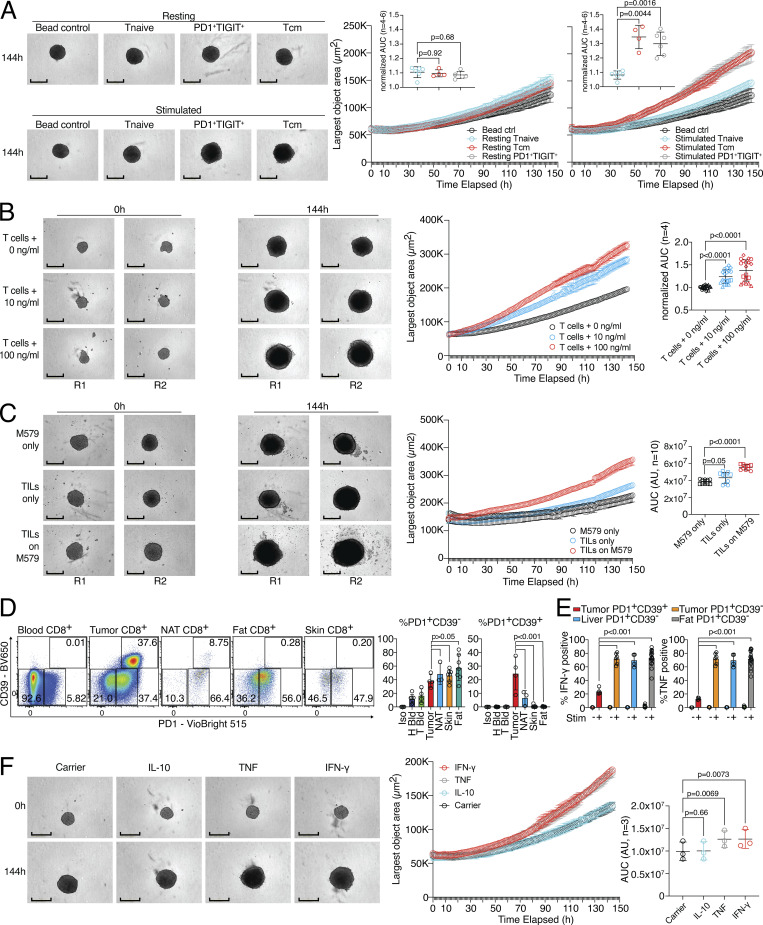
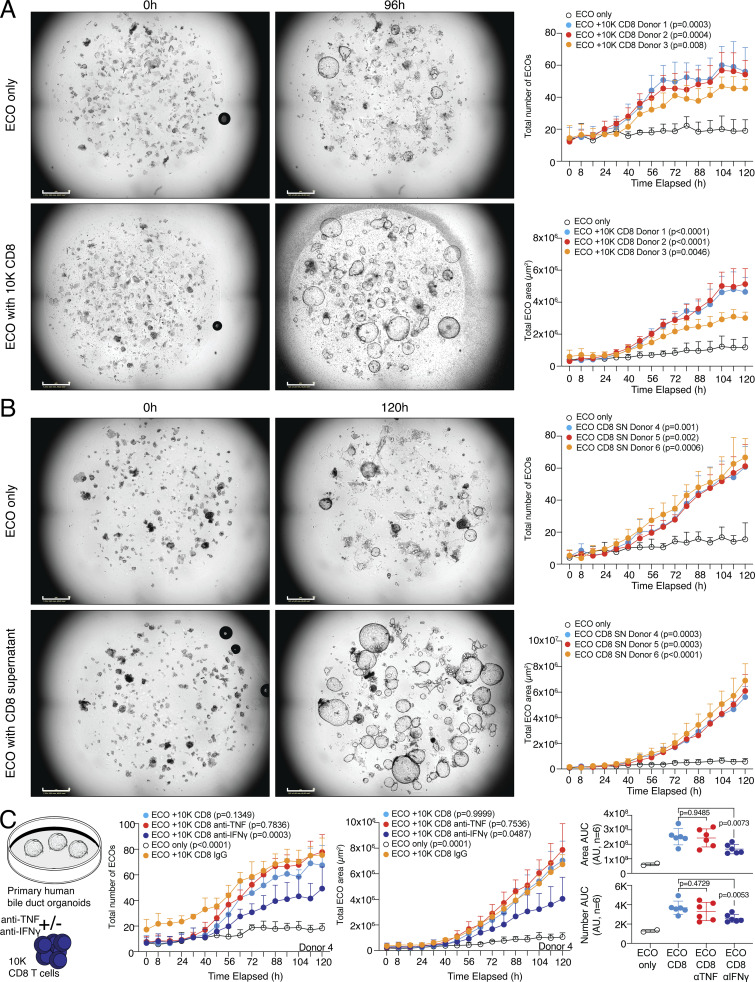
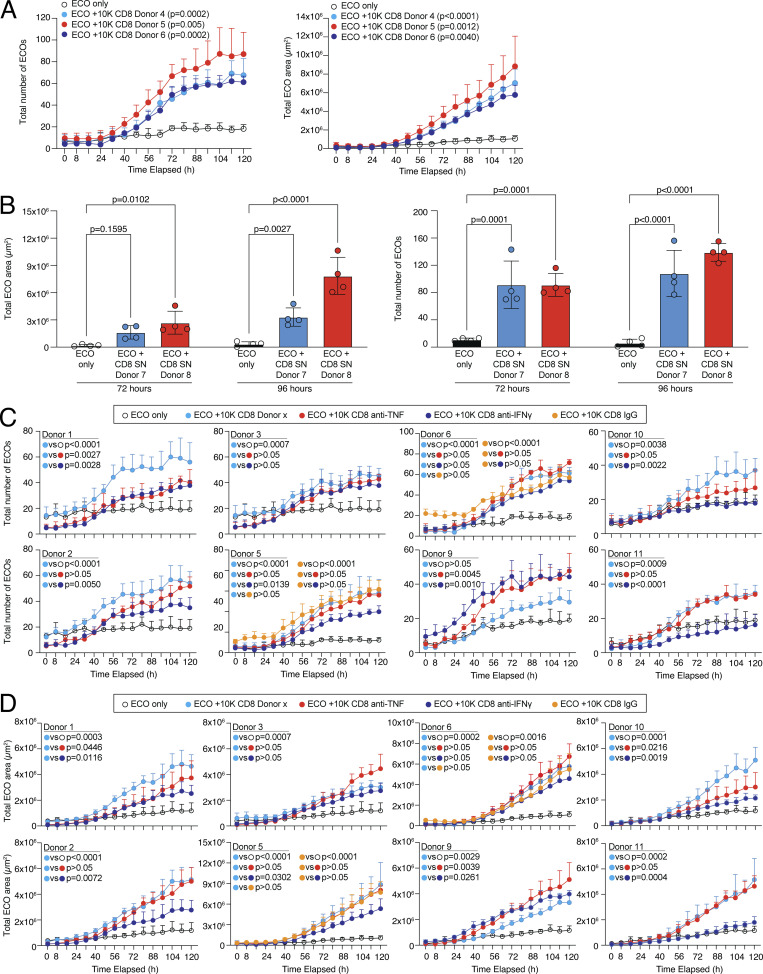
CD8 T lymphocytes are classically viewed as cytotoxic T cells. Whether human CD8 T cells can, in parallel, induce a tissue regeneration program is poorly understood. Here, antigen-specific assay systems revealed that human CD8 T cells not only mediated cytotoxicity but also promoted tissue remodeling. Activated CD8 T cells could produce the epidermal growth factor receptor (EGFR)-ligand amphiregulin (AREG) and sensitize epithelial cells for enhanced regeneration potential. Blocking the EGFR or the effector cytokines IFN-γ and TNF could inhibit tissue remodeling. This regenerative program enhanced tumor spheroid and stem cell-mediated organoid growth. Using single-cell gene expression analysis, we identified an AREG+, tissue-resident CD8 T cell population in skin and adipose tissue from patients undergoing abdominal wall or abdominoplasty surgery. These tissue-resident CD8 T cells showed a strong TCR clonal relation to blood PD1+TIGIT+ CD8 T cells with tissue remodeling abilities. These findings may help to understand the complex CD8 biology in tumors and could become relevant for the design of therapeutic T cell products.
© 2024 Delacher et al.
Conflict of interest statement
Disclosures: M. Delacher reported personal fees from Odyssey Therapeutics outside the submitted work. E.K. Geissler reported personal fees from Quell outside the submitted work. No other disclosures were reported.
Figures
References
-
- Bailey, S.R., Vatsa S., Larson R.C., Bouffard A.A., Scarfò I., Kann M.C., Berger T.R., Leick M.B., Wehrli M., Schmidts A., et al. . 2022. Blockade or deletion of IFNγ reduces macrophage activation without compromising CAR T-cell function in hematologic malignancies. Blood Cancer Discov. 3:136–153. 10.1158/2643-3230.BCD-21-0181 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
