

TET3 is a positive regulator of mitochondrial respiration in Neuro2A cells
- PMID: 38227585
- PMCID: PMC10790995
- DOI: 10.1371/journal.pone.0294187
TET3 is a positive regulator of mitochondrial respiration in Neuro2A cells
Erratum in
-
Correction: TET3 is a positive regulator of mitochondrial respiration in Neuro2A cells.PLoS One. 2024 Feb 29;19(2):e0300015. doi: 10.1371/journal.pone.0300015. eCollection 2024. PLoS One. 2024. PMID: 38422044 Free PMC article.
Abstract
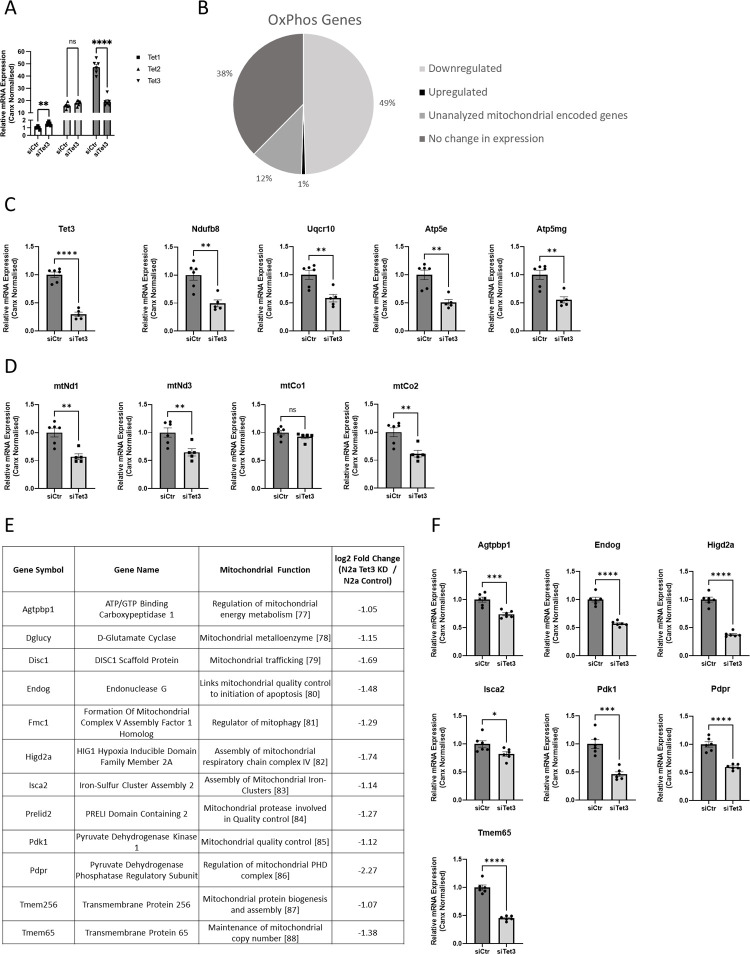
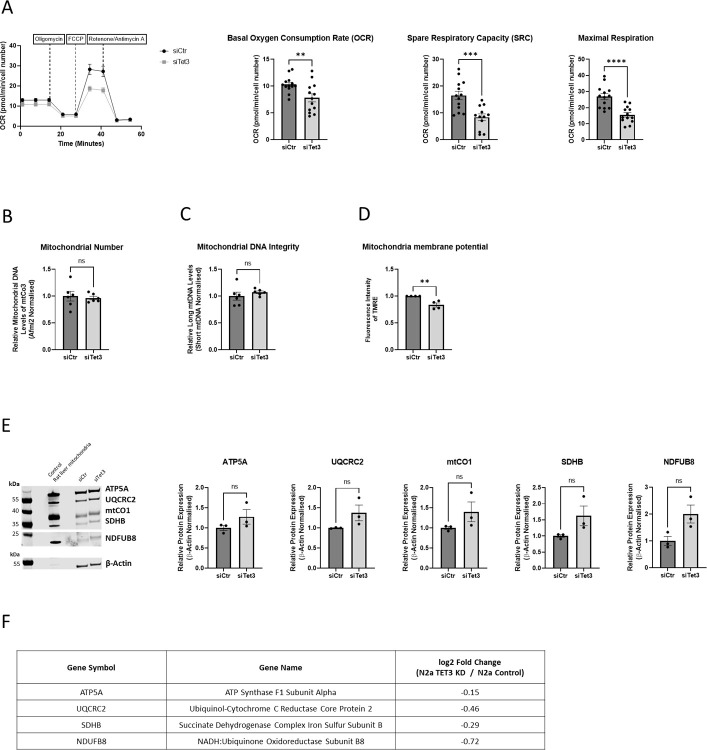
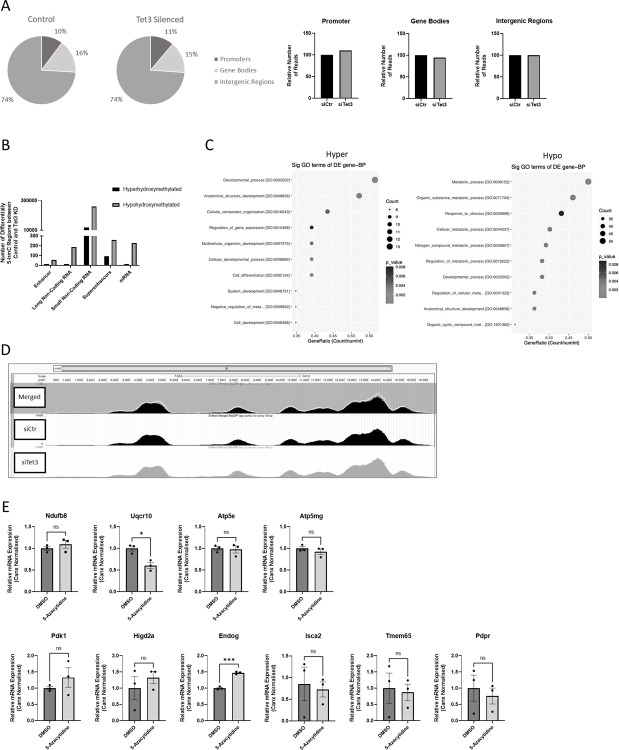
Ten-Eleven-Translocase (TET) enzymes contribute to the regulation of the methylome via successive oxidation of 5-methyl cytosine (5mC) to derivatives which can be actively removed by base-excision-repair (BER) mechanisms in the absence of cell division. This is particularly important in post-mitotic neurons where changes in DNA methylation are known to associate with changes in neural function. TET3, specifically, is a critical regulator of both neuronal differentiation in development and mediates dynamic changes in the methylome of adult neurons associated with cognitive function. While DNA methylation is understood to regulate transcription, little is known of the specific targets of TET3-dependent catalytic activity in neurons. We report the results of an unbiased transcriptome analysis of the neuroblastoma-derived cell line; Neuro2A, in which Tet3 was silenced. Oxidative phosphorylation (OxPhos) was identified as the most significantly down-regulated functional canonical pathway, and these findings were confirmed by measurements of oxygen consumption rate in the Seahorse bioenergetics analyser. The mRNA levels of both nuclear- and mitochondrial-encoded OxPhos genes were reduced by Tet3-silencing, but we found no evidence for differential (hydroxy)methylation deposition at these gene loci. However, the mRNA expression of genes known to be involved in mitochondrial quality control were also shown to be significantly downregulated in the absence of TET3. One of these genes; EndoG, was identified as a direct target of TET3-catalytic activity at non-CpG methylated sites within its gene body. Accordingly, we propose that aberrant mitochondrial homeostasis may contribute to the decrease in OxPhos, observed upon Tet3-downregulation in Neuro2A cells.
Copyright: © 2024 Kropf et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
