

γδ T cell antigen receptor polyspecificity enables T cell responses to a broad range of immune challenges
- PMID: 38227652
- PMCID: PMC10823224
- DOI: 10.1073/pnas.2315592121
γδ T cell antigen receptor polyspecificity enables T cell responses to a broad range of immune challenges
Abstract
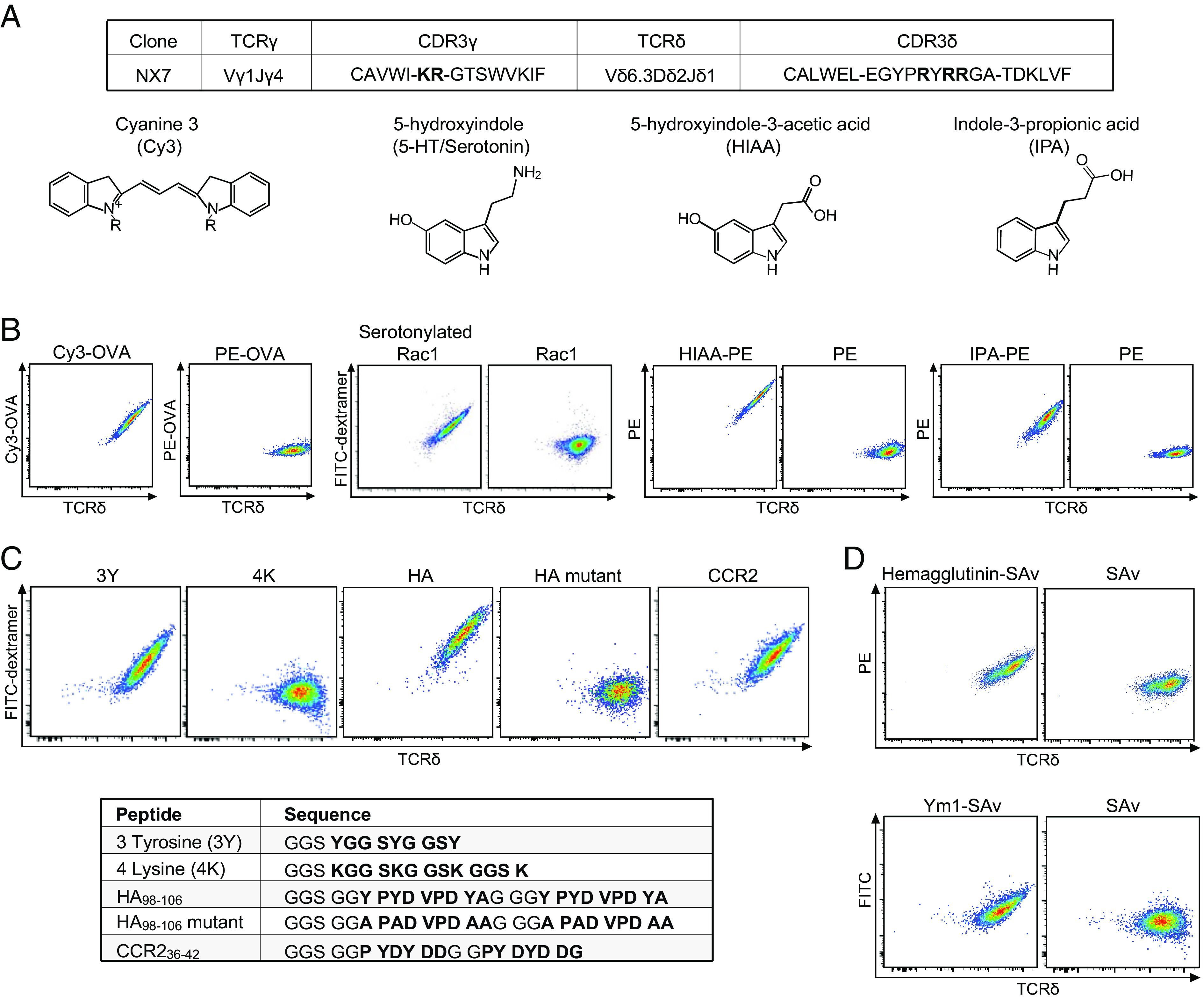
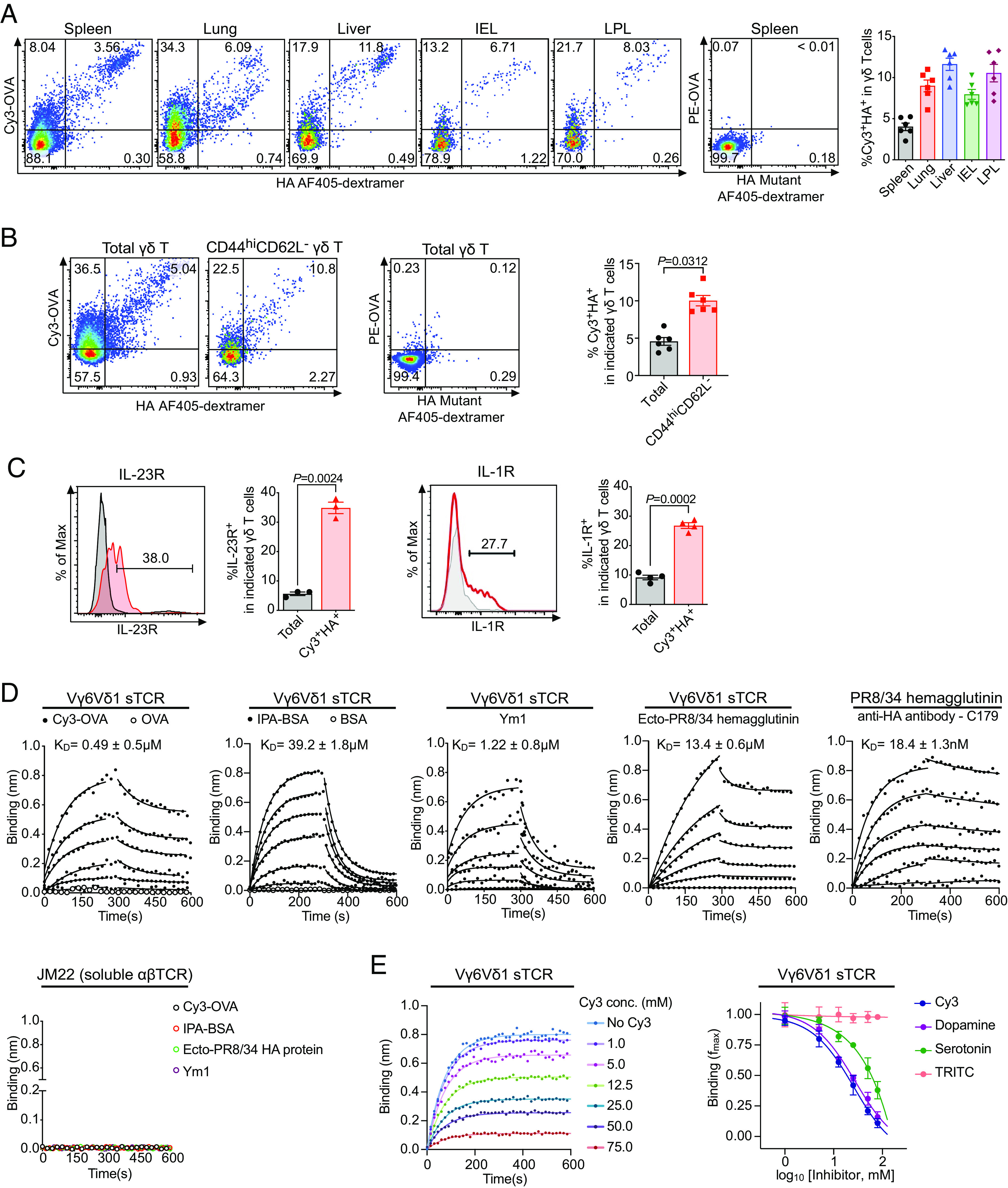
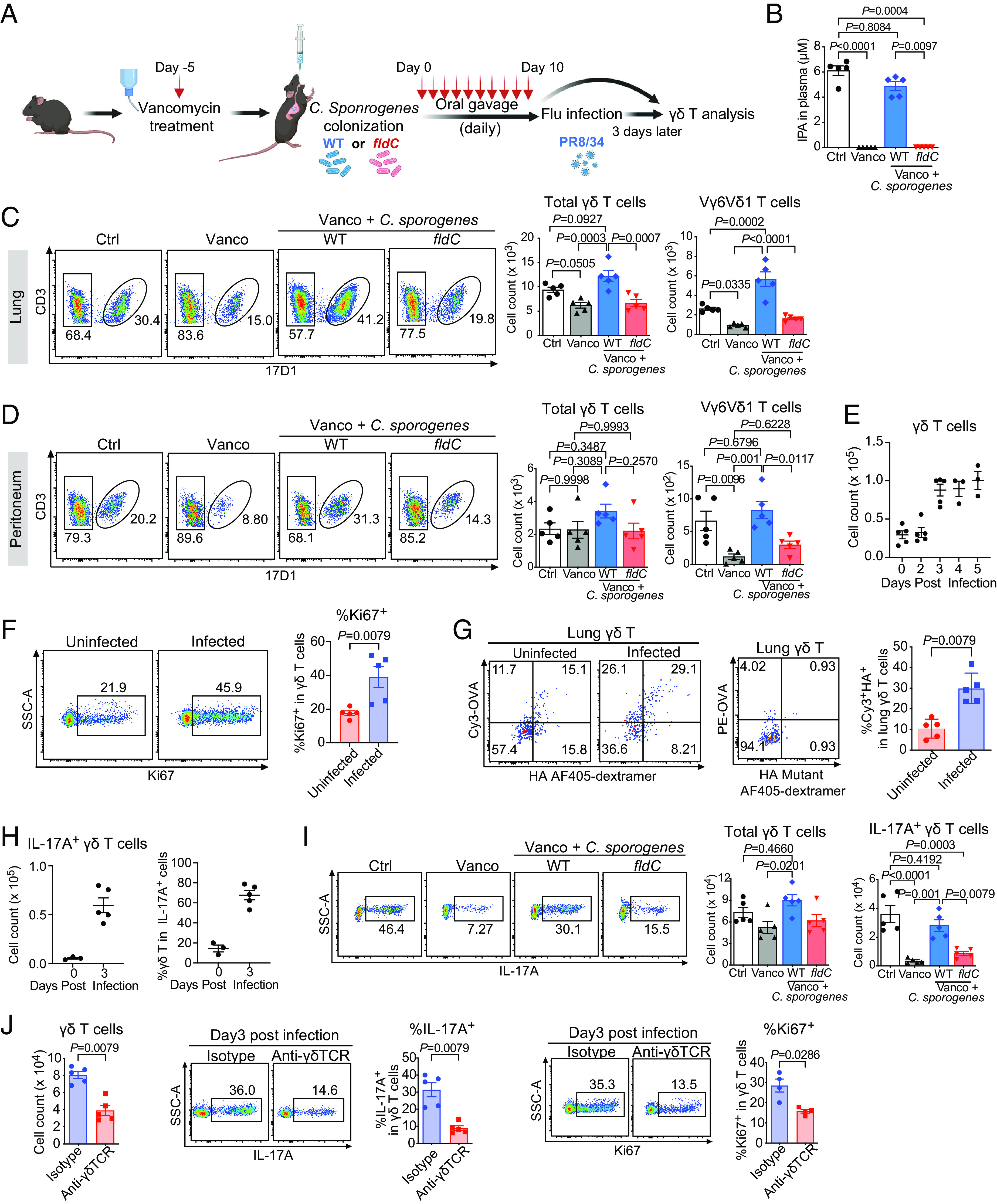
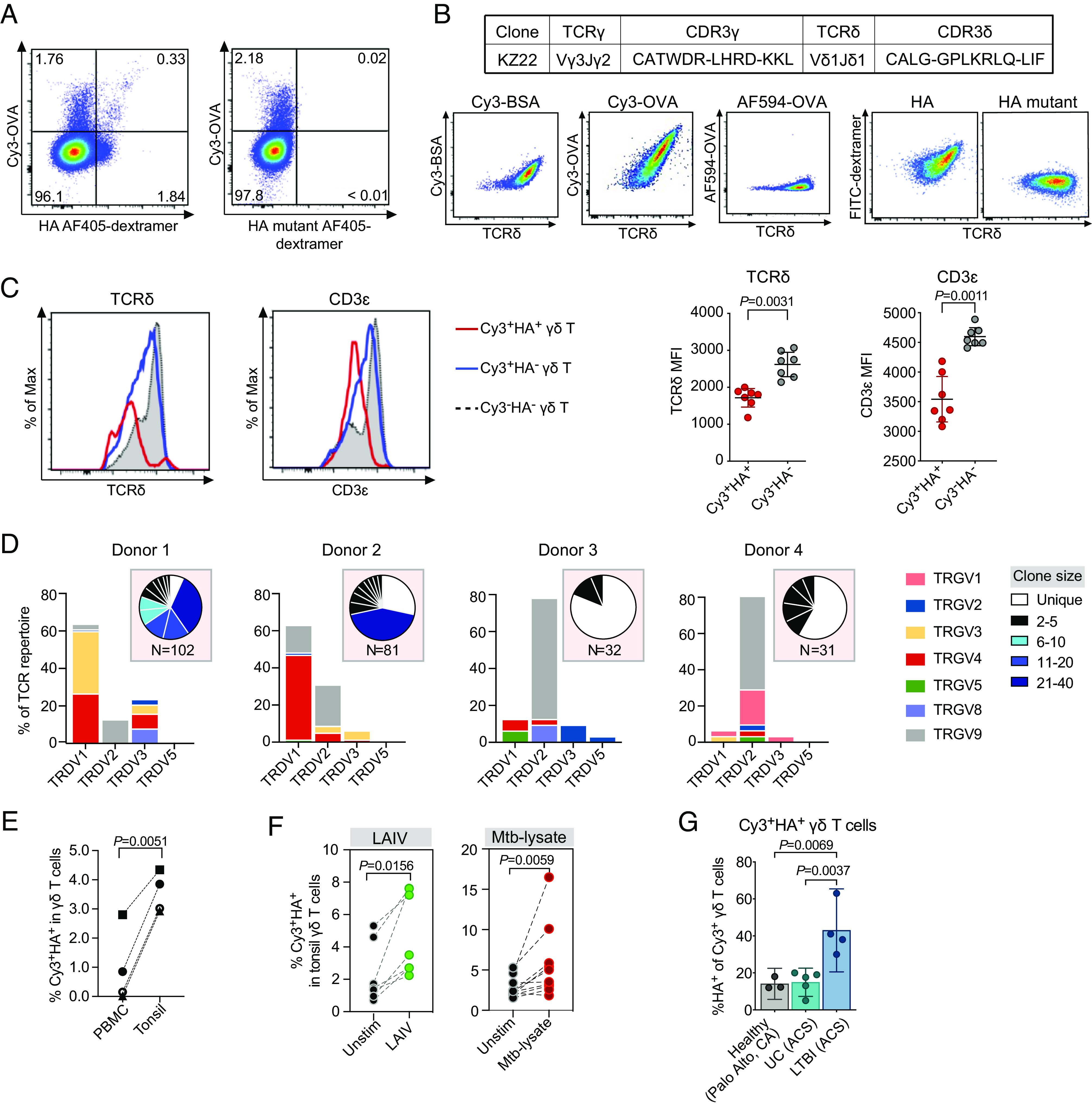
γδ T cells are essential for immune defense and modulating physiological processes. While they have the potential to recognize large numbers of antigens through somatic gene rearrangement, the antigens which trigger most γδ T cell response remain unidentified, and the role of antigen recognition in γδ T cell function is contentious. Here, we show that some γδ T cell receptors (TCRs) exhibit polyspecificity, recognizing multiple ligands of diverse molecular nature. These ligands include haptens, metabolites, neurotransmitters, posttranslational modifications, as well as peptides and proteins of microbial and host origin. Polyspecific γδ T cells are enriched among activated cells in naive mice and the responding population in infection. They express diverse TCR sequences, have different functional potentials, and include the innate-like γδ T cells, such as the major IL-17 responders in various pathological/physiological conditions. We demonstrate that encountering their antigenic microbiome metabolite maintains their homeostasis and functional response, indicating that their ability to recognize multiple ligands is essential for their function. Human γδ T cells with similar polyspecificity also respond to various immune challenges. This study demonstrates that polyspecificity is a prevalent feature of γδ T cell antigen recognition, which enables rapid and robust T cell responses to a wide range of challenges, highlighting a unique function of γδ T cells.
Keywords: antigen receptor poly-specificity; γδ T cell antigen recognition and response; γδ T cells.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
Comment in
-
Broad-spectrum defenders: γδ T cells take on a multitude of immune challenges.J Leukoc Biol. 2024 Apr 29;115(5):794-796. doi: 10.1093/jleuko/qiae044. J Leukoc Biol. 2024. PMID: 38411623
References
-
- Chien Y. H., Meyer C., Bonneville M., gammadelta T cells: First line of defense and beyond. Annu. Rev. Immunol. 32, 121–155 (2014). - PubMed
-
- Ribot J. C., Lopes N., Silva-Santos B., γδ T cells in tissue physiology and surveillance. Nat. Rev. Immunol. 21, 221–232 (2020). - PubMed
-
- Papotto P. H., Reinhardt A., Prinz I., Silva-Santos B., Innately versatile: Gammadelta17 T cells in inflammatory and autoimmune diseases. J. Autoimmun. 87, 26–37 (2018). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
