

Lack of affinity signature for germinal center cells that have initiated plasma cell differentiation
- PMID: 38228150
- PMCID: PMC10922795
- DOI: 10.1016/j.immuni.2023.12.010
Lack of affinity signature for germinal center cells that have initiated plasma cell differentiation
Abstract
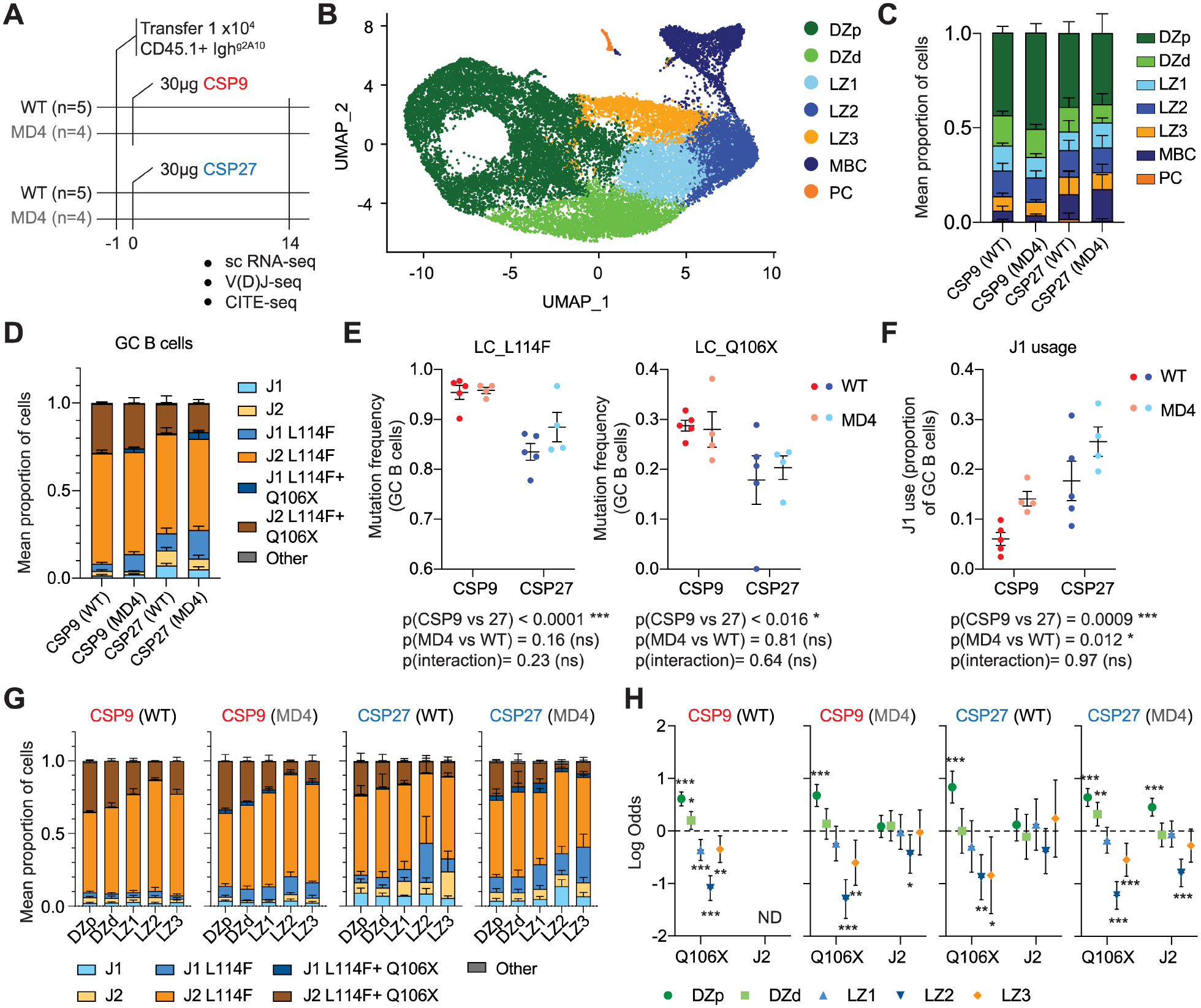
Long-lived plasma cells (PCs) secrete antibodies that can provide sustained immunity against infection. High-affinity cells are proposed to preferentially select into this compartment, potentiating the immune response. We used single-cell RNA-seq to track the germinal center (GC) development of Ighg2A10 B cells, specific for the Plasmodium falciparum circumsporozoite protein (PfCSP). Following immunization with Plasmodium sporozoites, we identified 3 populations of cells in the GC light zone (LZ). One LZ population expressed a gene signature associated with the initiation of PC differentiation and readily formed PCs in vitro. The estimated affinity of these pre-PC B cells was indistinguishable from that of LZ cells that remained in the GC. This remained true when high- or low-avidity recombinant PfCSP proteins were used as immunogens. These findings suggest that the initiation of PC development occurs via an affinity-independent process.
Keywords: B cells; Plasmodium; affinity maturation; germinal center; plasma cells; single-cell RNA-seq.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



References
-
- Jerne NK (1951). A study of avidity based on rabbit skin responses to diphtheria toxin-antitoxin mixtures. Acta Pathol Microbiol Scand Suppl (1926) 87, 1–183. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
