

Biomimetic Synthesis and Chemical Proteomics Reveal the Mechanism of Action and Functional Targets of Phloroglucinol Meroterpenoids
- PMID: 38230968
- PMCID: PMC11000255
- DOI: 10.1021/jacs.3c10741
Biomimetic Synthesis and Chemical Proteomics Reveal the Mechanism of Action and Functional Targets of Phloroglucinol Meroterpenoids
Abstract
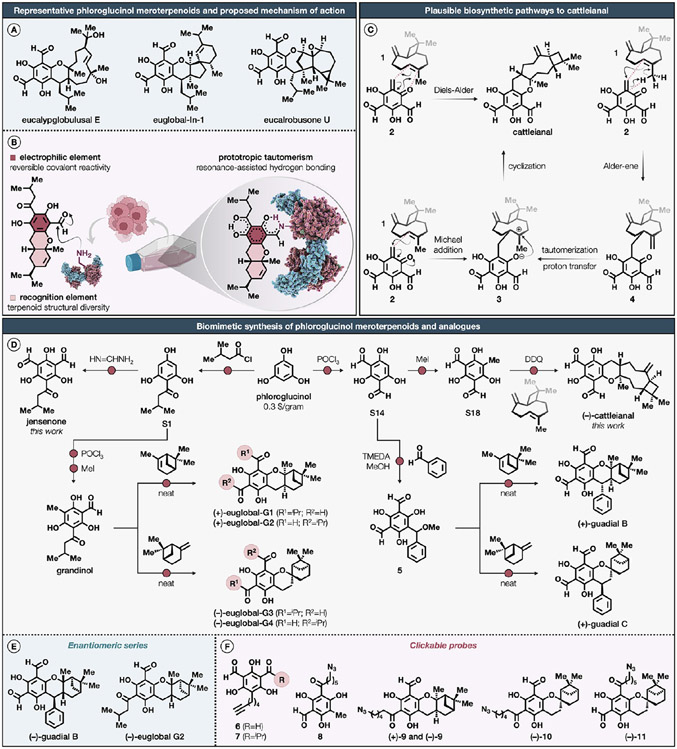
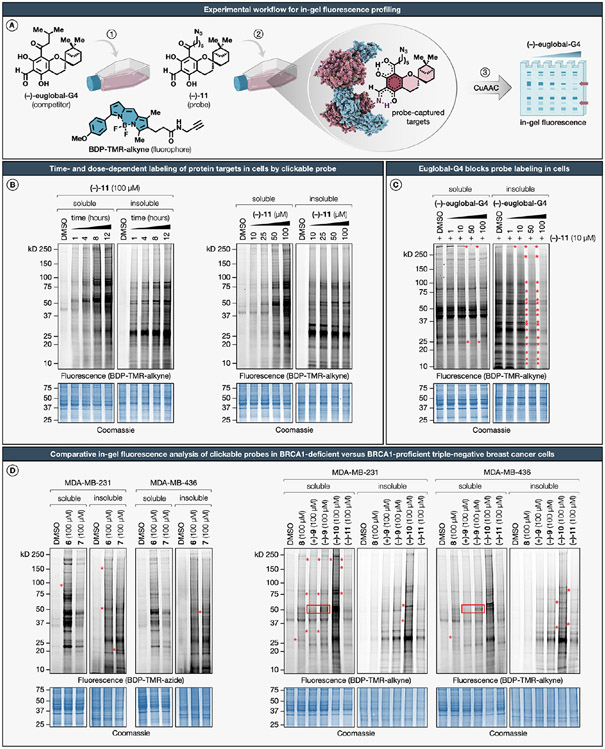
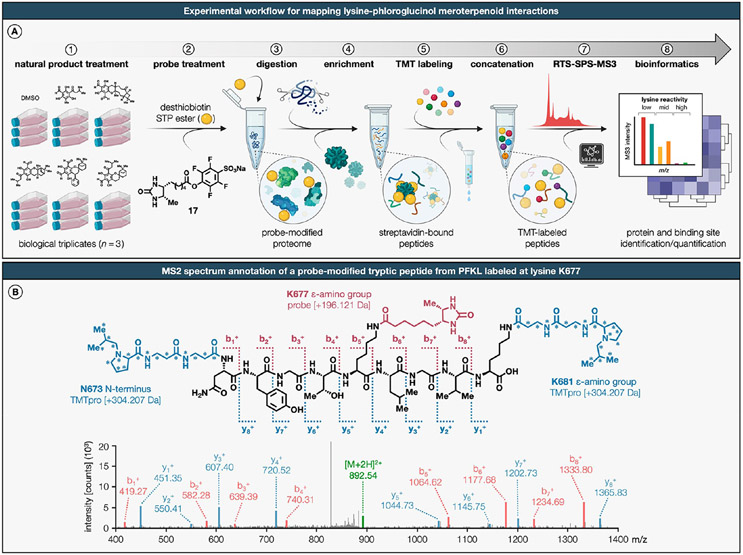
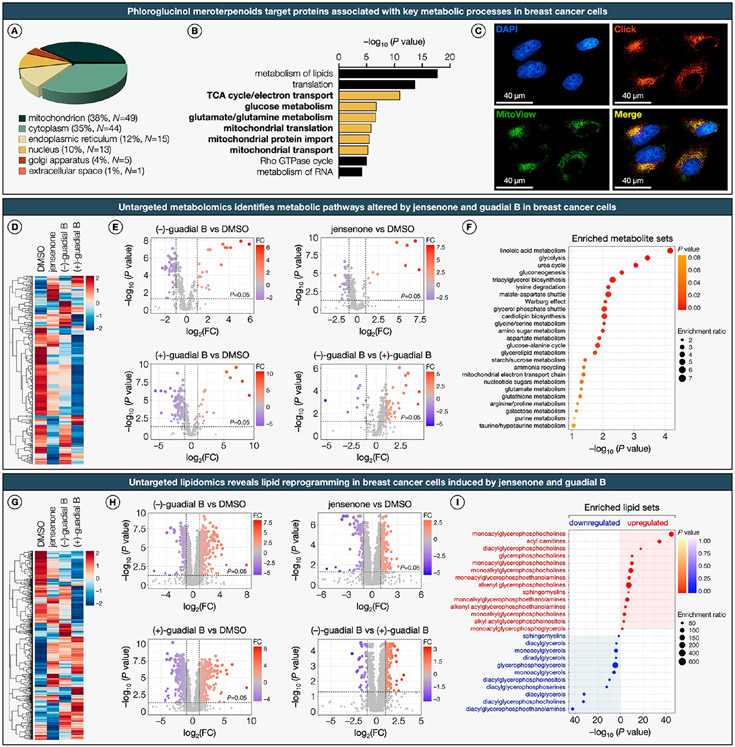
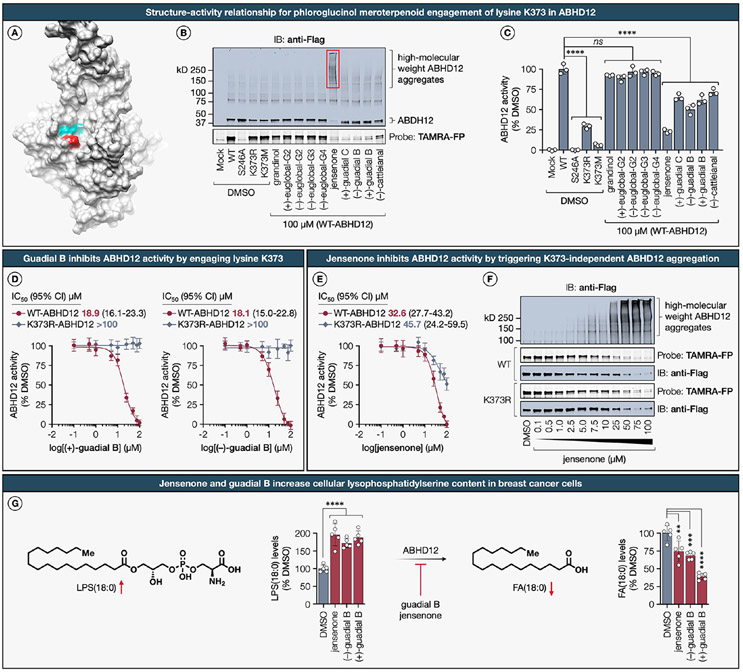
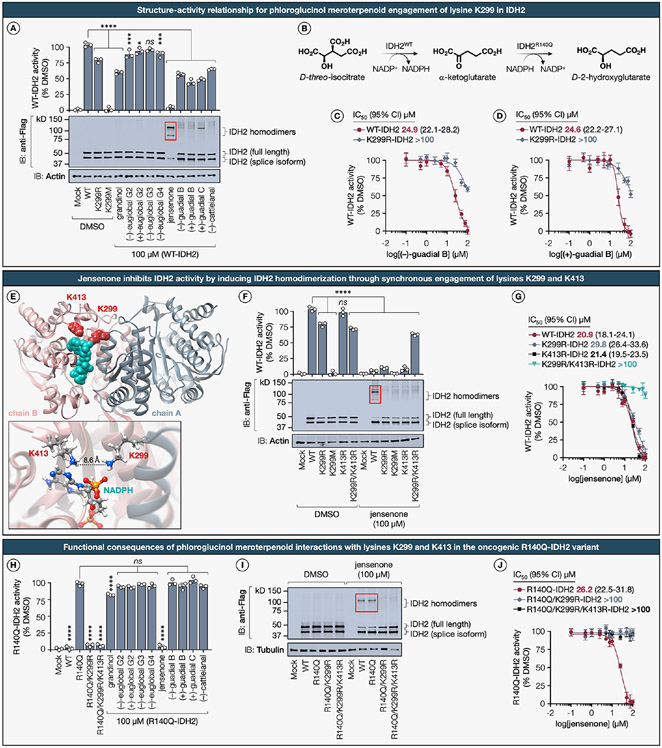
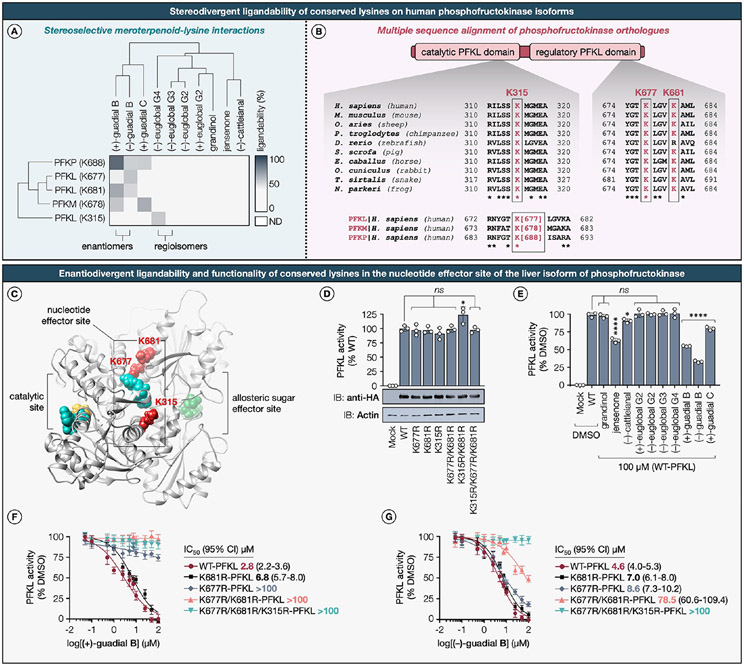
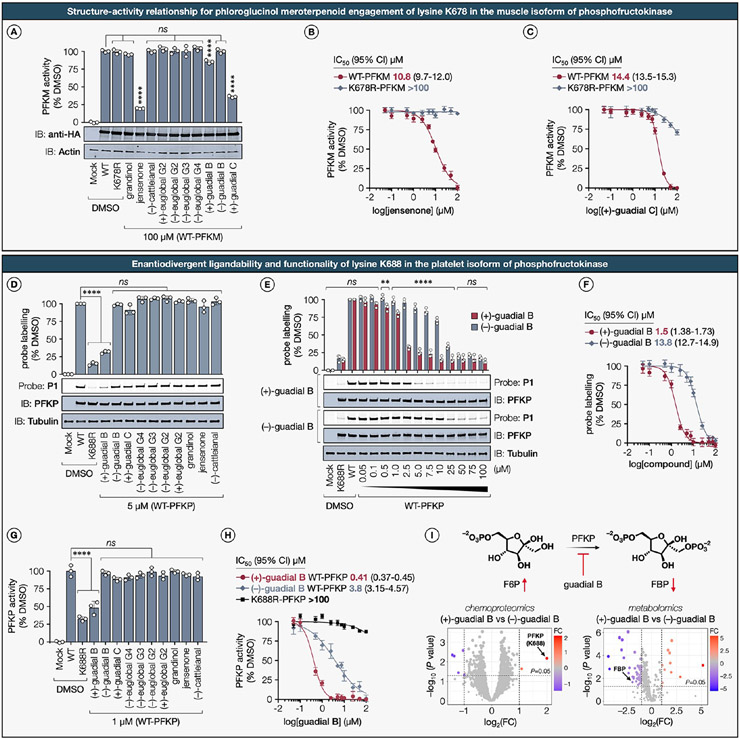
Natural products perennially serve as prolific sources of drug leads and chemical probes, fueling the development of numerous therapeutics. Despite their scarcity, natural products that modulate protein function through covalent interactions with lysine residues hold immense potential to unlock new therapeutic interventions and advance our understanding of the biological processes governed by these modifications. Phloroglucinol meroterpenoids constitute one of the most expansive classes of natural products, displaying a plethora of biological activities. However, their mechanism of action and cellular targets have, until now, remained elusive. In this study, we detail the concise biomimetic synthesis, computational mechanistic insights, physicochemical attributes, kinetic parameters, molecular mechanism of action, and functional cellular targets of several phloroglucinol meroterpenoids. We harness synthetic clickable analogues of natural products to probe their disparate proteome-wide reactivity and subcellular localization through in-gel fluorescence scanning and cell imaging. By implementing sample multiplexing and a redesigned lysine-targeting probe, we streamline a quantitative activity-based protein profiling, enabling the direct mapping of global reactivity and ligandability of proteinaceous lysines in human cells. Leveraging this framework, we identify numerous lysine-meroterpenoid interactions in breast cancer cells at tractable protein sites across diverse structural and functional classes, including those historically deemed undruggable. We validate that phloroglucinol meroterpenoids perturb biochemical functions through stereoselective and site-specific modification of lysines in proteins vital for breast cancer metabolism, including lipid signaling, mitochondrial respiration, and glycolysis. These findings underscore the broad potential of phloroglucinol meroterpenoids for targeting functional lysines in the human proteome.
Figures

Similar articles
-
A proteome-wide atlas of lysine-reactive chemistry.Nat Chem. 2021 Nov;13(11):1081-1092. doi: 10.1038/s41557-021-00765-4. Epub 2021 Sep 9. Nat Chem. 2021. PMID: 34504315 Free PMC article.
-
Global profiling of lysine reactivity and ligandability in the human proteome.Nat Chem. 2017 Dec;9(12):1181-1190. doi: 10.1038/nchem.2826. Epub 2017 Jul 31. Nat Chem. 2017. PMID: 29168484 Free PMC article.
-
[Advances in applications of activity-based chemical probes in the characterization of amino acid reactivities].Se Pu. 2023 Jan;41(1):14-23. doi: 10.3724/SP.J.1123.2022.05013. Se Pu. 2023. PMID: 36633073 Free PMC article. Review. Chinese.
-
Global Profiling Lysine Reactivity and Ligandability with Oxidant-Triggered Bioconjugation Chemistry.Angew Chem Int Ed Engl. 2025 Feb 3;64(6):e202418473. doi: 10.1002/anie.202418473. Epub 2024 Nov 29. Angew Chem Int Ed Engl. 2025. PMID: 39543955
-
Peculiarities of meroterpenoids and their bioproduction.Appl Microbiol Biotechnol. 2021 May;105(10):3987-4003. doi: 10.1007/s00253-021-11312-z. Epub 2021 May 3. Appl Microbiol Biotechnol. 2021. PMID: 33937926 Review.
Cited by
-
Activity-Based Acylome Profiling with N-(Cyanomethyl)-N-(phenylsulfonyl)amides for Targeted Lysine Acylation and Post-Translational Control of Protein Function in Cells.J Am Chem Soc. 2024 Oct 9;146(40):27622-27643. doi: 10.1021/jacs.4c09073. Epub 2024 Sep 30. J Am Chem Soc. 2024. PMID: 39348182 Free PMC article.
-
RNA G-quadruplexes: emerging regulators of gene expression and therapeutic targets.Funct Integr Genomics. 2025 Jul 3;25(1):143. doi: 10.1007/s10142-025-01656-4. Funct Integr Genomics. 2025. PMID: 40608121 Review.
-
Chemical tools to expand the ligandable proteome: Diversity-oriented synthesis-based photoreactive stereoprobes.Cell Chem Biol. 2024 Dec 19;31(12):2138-2155.e32. doi: 10.1016/j.chembiol.2024.10.005. Epub 2024 Nov 14. Cell Chem Biol. 2024. PMID: 39547236
-
Advancing Covalent Ligand and Drug Discovery beyond Cysteine.Chem Rev. 2025 Jul 23;125(14):6653-6684. doi: 10.1021/acs.chemrev.5c00001. Epub 2025 May 22. Chem Rev. 2025. PMID: 40404146 Free PMC article. Review.
-
Functionalizing tandem mass tags for streamlining click-based quantitative chemoproteomics.Commun Chem. 2024 Apr 10;7(1):80. doi: 10.1038/s42004-024-01162-x. Commun Chem. 2024. PMID: 38600184 Free PMC article.
References
-
- Hopkins AL; Groom CR The druggable genome. Nat. Rev. Drug Discovery 2002, 1 (9), 727–730. - PubMed
-
- Macarron R; Banks MN; Bojanic D; Burns DJ; Cirovic DA; Garyantes T; Green DVS; Hertzberg RP; Janzen WP; Paslay JW; et al. Impact of high-throughput screening in biomedical research. Nat. Rev. Drug Discovery 2011, 10 (3), 188–195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
