

TMEM106B core deposition associates with TDP-43 pathology and is increased in risk SNP carriers for frontotemporal dementia
- PMID: 38232138
- PMCID: PMC10841341
- DOI: 10.1126/scitranslmed.adf9735
TMEM106B core deposition associates with TDP-43 pathology and is increased in risk SNP carriers for frontotemporal dementia
Abstract
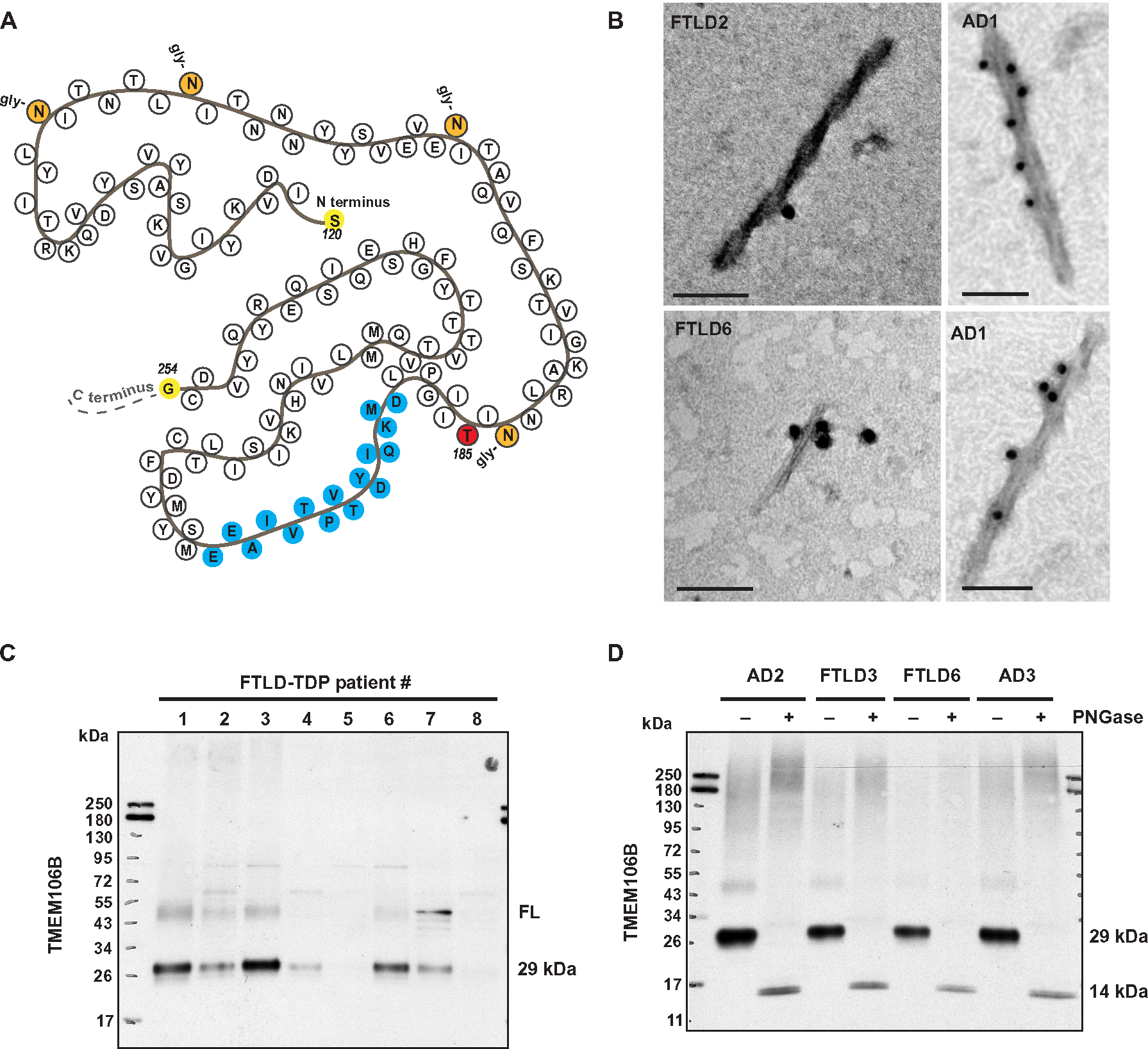
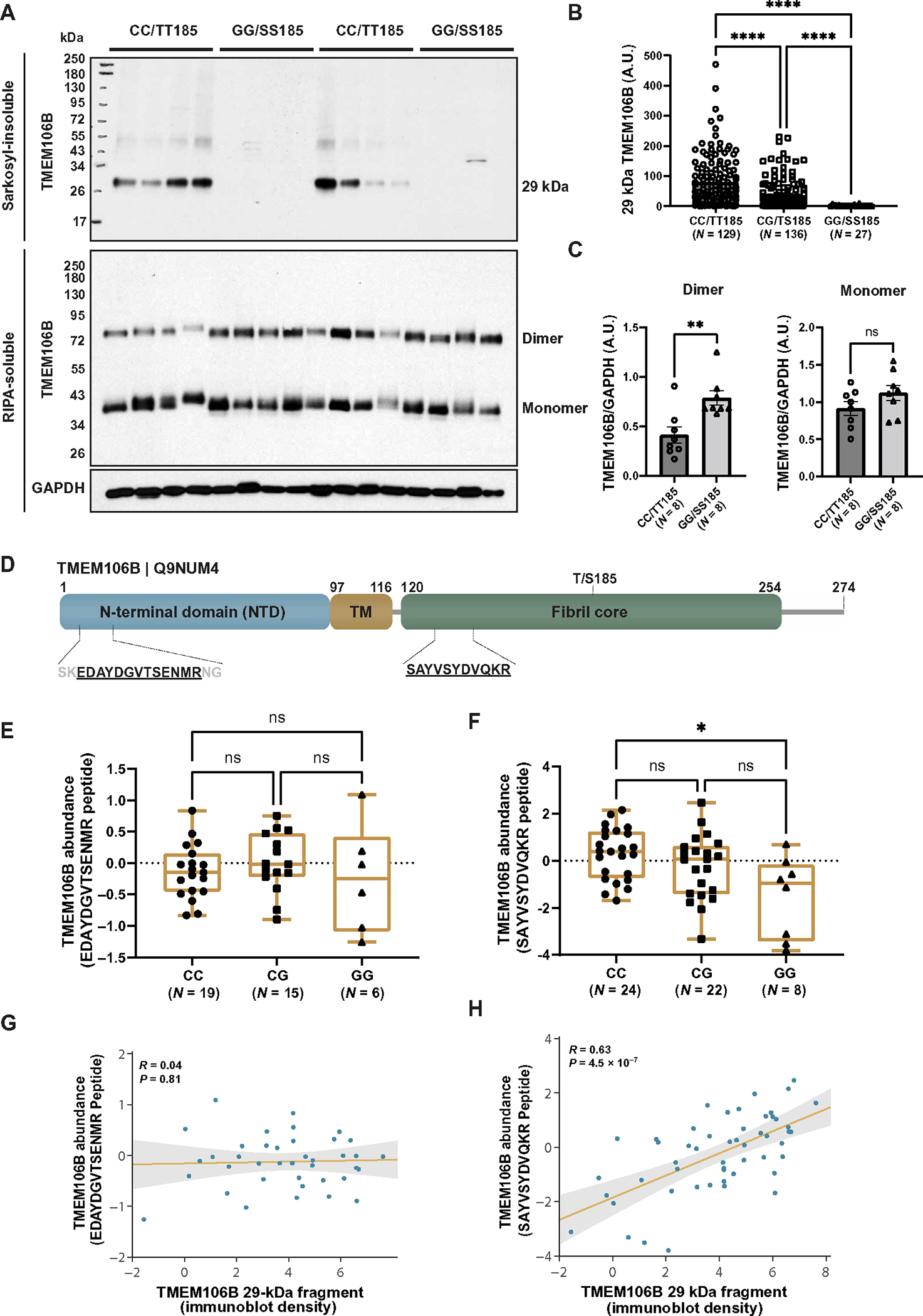
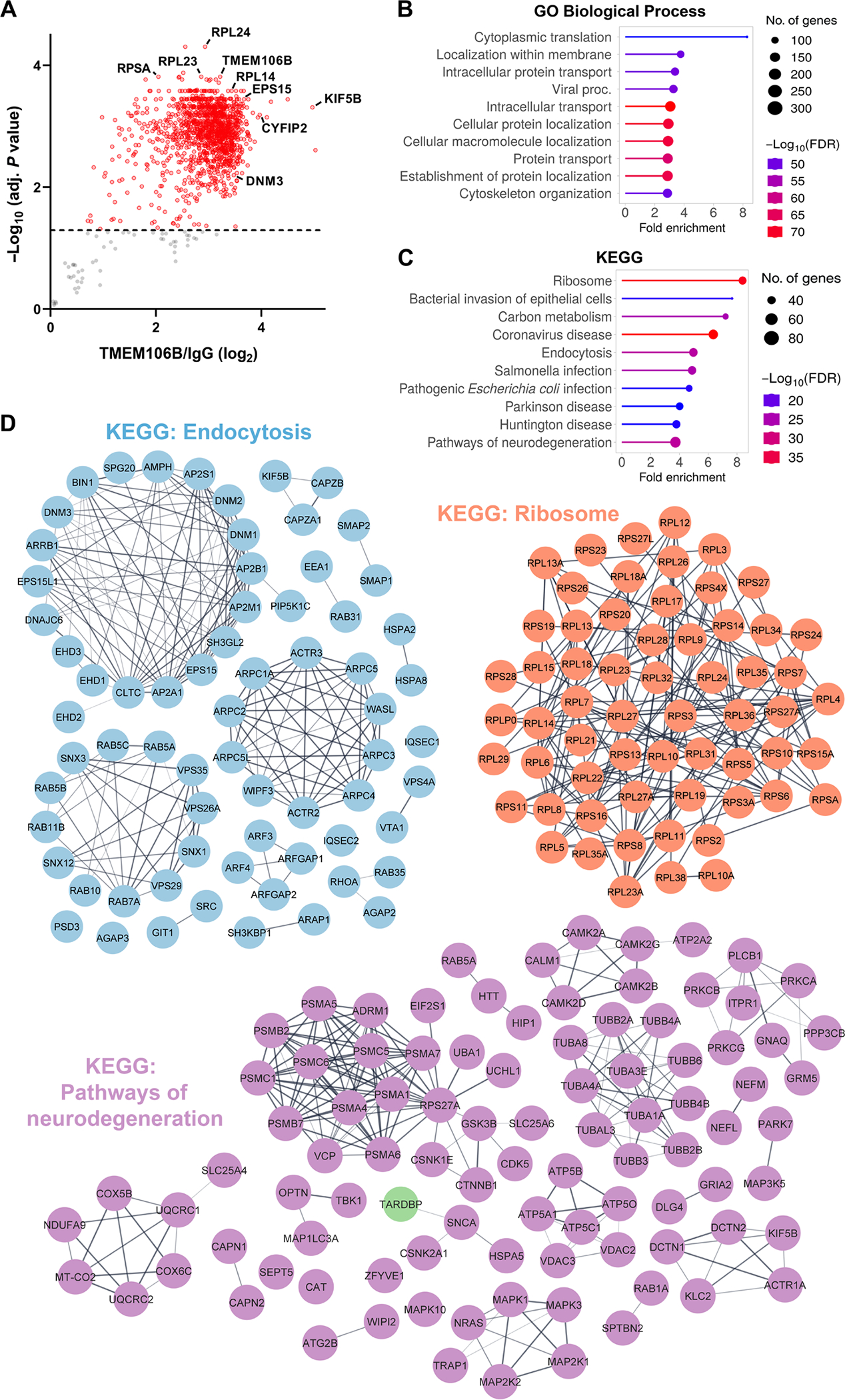
Genetic variation at the transmembrane protein 106B gene (TMEM106B) has been linked to risk of frontotemporal lobar degeneration with TDP-43 inclusions (FTLD-TDP) through an unknown mechanism. We found that presence of the TMEM106B rs3173615 protective genotype was associated with longer survival after symptom onset in a postmortem FTLD-TDP cohort, suggesting a slower disease course. The seminal discovery that filaments derived from TMEM106B is a common feature in aging and, across a range of neurodegenerative disorders, suggests that genetic variants in TMEM106B could modulate disease risk and progression through modulating TMEM106B aggregation. To explore this possibility and assess the pathological relevance of TMEM106B accumulation, we generated a new antibody targeting the TMEM106B filament core sequence. Analysis of postmortem samples revealed that the TMEM106B rs3173615 risk allele was associated with higher TMEM106B core accumulation in patients with FTLD-TDP. In contrast, minimal TMEM106B core deposition was detected in carriers of the protective allele. Although the abundance of monomeric full-length TMEM106B was unchanged, carriers of the protective genotype exhibited an increase in dimeric full-length TMEM106B. Increased TMEM106B core deposition was also associated with enhanced TDP-43 dysfunction, and interactome data suggested a role for TMEM106B core filaments in impaired RNA transport, local translation, and endolysosomal function in FTLD-TDP. Overall, these findings suggest that prevention of TMEM106B core accumulation is central to the mechanism by which the TMEM106B protective haplotype reduces disease risk and slows progression.
Conflict of interest statement
Figures
Comment in
-
Risk and progression of frontotemporal dementia in carriers of the TMEM106B protective genotype and its relationship with TDP-43 pathology.Ann Transl Med. 2024 Dec 24;12(6):119. doi: 10.21037/atm-24-104. Epub 2024 Dec 2. Ann Transl Med. 2024. PMID: 39817242 Free PMC article. No abstract available.
References
-
- Neary D, Snowden JS, Gustafson L, Passant U, Stuss D, Black S, Freedman M, Kertesz A, Robert PH, Albert M, Boone K, Miller BL, Cummings J, Benson DF, Frontotemporal lobar degeneration : A consensus on clinical diagnostic criteria. Neurology 51, 1546–1554 (1998). - PubMed
-
- Arai T, Hasegawa M, Akiyama H, Ikeda K, Nonaka T, Mori H, Mann D, Tsuchiya K, Yoshida M, Hashizume Y, Oda T, TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem. Biophys. Res. Commun. 351, 602–611 (2006). - PubMed
-
- Neumann M, Kwong LK, Truax AC, Vanmassenhove B, Kretzschmar HA, Van Deerlin VM, Clark CM, Grossman M, Miller BL, Trojanowski JQ, Lee VM-Y, TDP-43-positive white matter pathology in frontotemporal lobar degeneration with ubiquitin-positive inclusions. J. Neuropathol. Exp. Neurol. 66, 177–183 (2007). - PubMed
-
- Gallagher MD, Suh E, Grossman M, Elman L, McCluskey L, Van Swieten JC, Al-Sarraj S, Neumann M, Gelpi E, Ghetti B, Rohrer JD, Halliday G, Van Broeckhoven C, Seilhean D, Shaw PJ, Frosch MP, Alafuzoff I, Antonell A, Bogdanovic N, Brooks W, Cairns NJ, Cooper-Knock J, Cotman C, Cras P, Cruts M, De Deyn PP, DeCarli C, Dobson-Stone C, Engelborghs S, Fox N, Galasko D, Gearing M, Gijselinck I, Grafman J, Hartikainen P, Hatanpaa KJ, Highley JR, Hodges J, Hulette C, Ince PG, Jin LW, Kirby J, Kofler J, Kril J, Kwok JB, Levey A, Lieberman A, Llado A, Martin JJ, Masliah E, McDermott CJ, McKee A, McLean C, Mead S, Miller CA, Miller J, Munoz DG, Murrell J, Paulson H, Piguet O, Rossor M, Sanchez-Valle R, Sano M, Schneider J, Silbert LC, Spina S, van der Zee J, Van Langenhove T, Warren J, Wharton SB, White CL 3rd, Woltjer RL, Trojanowski JQ, Lee VM, Van Deerlin V, Chen-Plotkin AS, TMEM106B is a genetic modifier of frontotemporal lobar degeneration with C9orf72 hexanucleotide repeat expansions. Acta Neuropathol. 127, 407–418 (2014). - PMC - PubMed
-
- van Blitterswijk M, Mullen B, Nicholson AM, Bieniek KF, Heckman MG, Baker MC, DeJesus-Hernandez M, Finch NA, Brown PH, Murray ME, Hsiung GY, Stewart H, Karydas AM, Finger E, Kertesz A, Bigio EH, Weintraub S, Mesulam M, Hatanpaa KJ, White CL 3rd, Strong MJ, Beach TG, Wszolek ZK, Lippa C, Caselli R, Petrucelli L, Josephs KA, Parisi JE, Knopman DS, Petersen RC, Mackenzie IR, Seeley WW, Grinberg LT, Miller BL, Boylan KB, Graff-Radford NR, Boeve BF, Dickson DW, Rademakers R, TMEM106B protects C9ORF72 expansion carriers against frontotemporal dementia. Acta Neuropathol. 127, 397–406 (2014). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AG063780/AG/NIA NIH HHS/United States
- R01 NS120992/NS/NINDS NIH HHS/United States
- U01 NS110438/NS/NINDS NIH HHS/United States
- RF1 AG062077/AG/NIA NIH HHS/United States
- U01 AG061357/AG/NIA NIH HHS/United States
- P01 AG019724/AG/NIA NIH HHS/United States
- R21 NS127331/NS/NINDS NIH HHS/United States
- P01 NS084974/NS/NINDS NIH HHS/United States
- U54 NS123743/NS/NINDS NIH HHS/United States
- RF1 NS120992/NS/NINDS NIH HHS/United States
- R01 NS117461/NS/NINDS NIH HHS/United States
- R35 NS097273/NS/NINDS NIH HHS/United States
- R01 AG071513/AG/NIA NIH HHS/United States
- RF1 AG062171/AG/NIA NIH HHS/United States
- R01 AG065219/AG/NIA NIH HHS/United States
- U19 AG063911/AG/NIA NIH HHS/United States
- F30 AG085984/AG/NIA NIH HHS/United States
- P30 AG062677/AG/NIA NIH HHS/United States
- R01 AG037491/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
