

Hypercapnia Causes Injury of the Cerebral Cortex and Cognitive Deficits in Newborn Piglets
- PMID: 38233145
- PMCID: PMC10913040
- DOI: 10.1523/ENEURO.0268-23.2023
Hypercapnia Causes Injury of the Cerebral Cortex and Cognitive Deficits in Newborn Piglets
Abstract
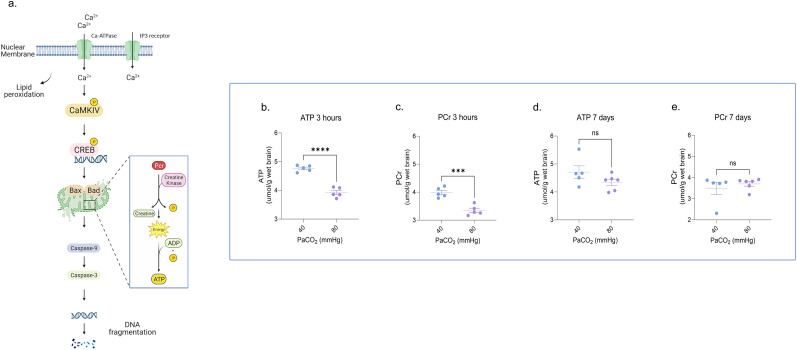
In critically ill newborns, exposure to hypercapnia (HC) is common and often accepted in neonatal intensive care units to prevent severe lung injury. However, as a "safe" range of arterial partial pressure of carbon dioxide levels in neonates has not been established, the potential impact of HC on the neurodevelopmental outcomes in these newborns remains a matter of concern. Here, in a newborn Yorkshire piglet model of either sex, we show that acute exposure to HC induced persistent cortical neuronal injury, associated cognitive and learning deficits, and long-term suppression of cortical electroencephalogram frequencies. HC induced a transient energy failure in cortical neurons, a persistent dysregulation of calcium-dependent proapoptotic signaling in the cerebral cortex, and activation of the apoptotic cascade, leading to nuclear deoxyribonucleic acid fragmentation. While neither 1 h of HC nor the rapid normalization of HC was associated with changes in cortical bioenergetics, rapid resuscitation resulted in a delayed onset of synaptosomal membrane lipid peroxidation, suggesting a dissociation between energy failure and the occurrence of synaptosomal lipid peroxidation. Even short durations of HC triggered biochemical responses at the subcellular level of the cortical neurons resulting in altered cortical activity and impaired neurobehavior. The deleterious effects of HC on the developing brain should be carefully considered as crucial elements of clinical decisions in the neonatal intensive care unit.
Keywords: cortex; hypercapnia; hypercarbia; neonatal; permissive; piglet.
Copyright © 2024 Fritz et al.
Figures




Similar articles
-
Hypercapnia-induced modifications of neuronal function in the cerebral cortex of newborn piglets.Pediatr Res. 2005 Feb;57(2):299-304. doi: 10.1203/01.PDR.0000148718.47137.9B. Epub 2004 Dec 7. Pediatr Res. 2005. PMID: 15585683
-
Hypercapnia and hypocapnia in neonates.World J Pediatr. 2008 Aug;4(3):192-6. doi: 10.1007/s12519-008-0035-5. World J Pediatr. 2008. PMID: 18822927 Review.
-
Computational analysis of cortical neuronal excitotoxicity in a large animal model of neonatal brain injury.J Neurodev Disord. 2022 Mar 29;14(1):26. doi: 10.1186/s11689-022-09431-3. J Neurodev Disord. 2022. PMID: 35351004 Free PMC article.
-
Hypoxia-induced Bax and Bcl-2 protein expression, caspase-9 activation, DNA fragmentation, and lipid peroxidation in mitochondria of the cerebral cortex of newborn piglets: the role of nitric oxide.Neuroscience. 2006 Sep 1;141(3):1339-49. doi: 10.1016/j.neuroscience.2006.05.005. Epub 2006 Jun 13. Neuroscience. 2006. PMID: 16777344
-
Carbon dioxide in the critically ill: too much or too little of a good thing?Respir Care. 2014 Oct;59(10):1597-605. doi: 10.4187/respcare.03405. Respir Care. 2014. PMID: 25261559 Review.
Cited by
-
Investigation of Suffocation Mechanisms in the Infant Sleep Environment Using a Mechanical Breathing Model Simulation.Cureus. 2025 Feb 11;17(2):e78852. doi: 10.7759/cureus.78852. eCollection 2025 Feb. Cureus. 2025. PMID: 40084315 Free PMC article.
-
The shifting landscape of the preterm brain.Neuron. 2025 Jul 9;113(13):2042-2064. doi: 10.1016/j.neuron.2025.03.024. Epub 2025 Apr 15. Neuron. 2025. PMID: 40239653 Free PMC article. Review.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources