

From structural polymorphism to structural metamorphosis of the coat protein of flexuous filamentous potato virus Y
- PMID: 38233506
- PMCID: PMC10794713
- DOI: 10.1038/s42004-024-01100-x
From structural polymorphism to structural metamorphosis of the coat protein of flexuous filamentous potato virus Y
Abstract
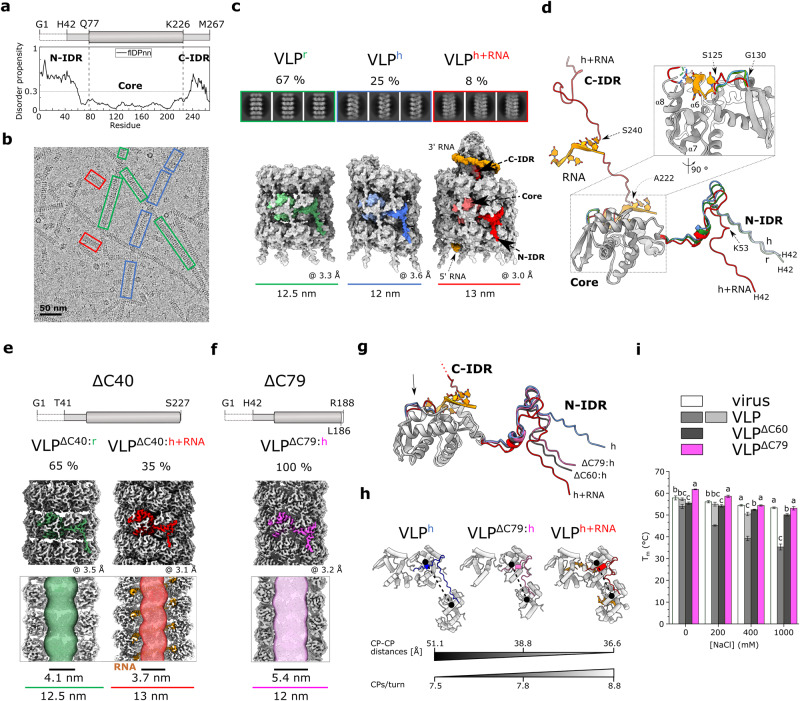
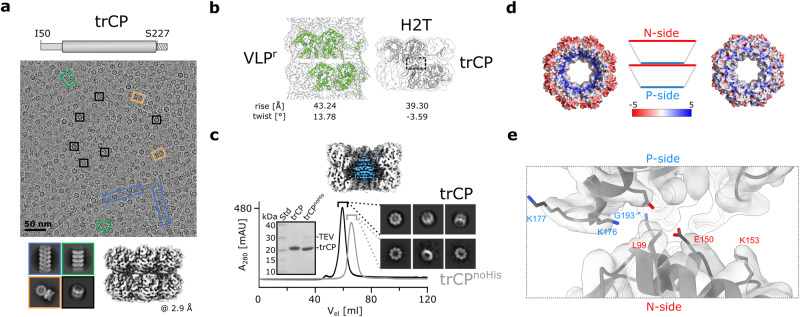
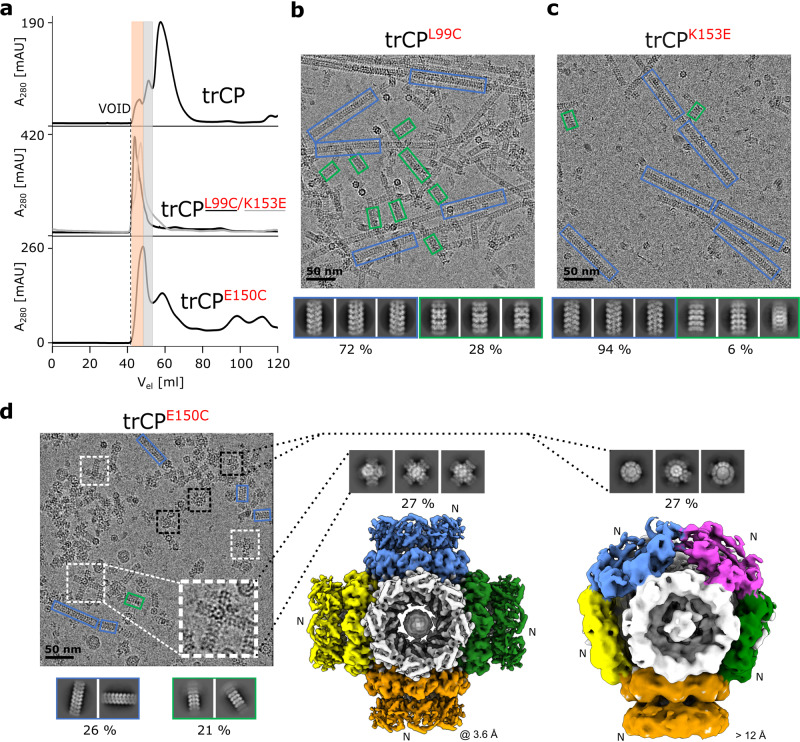
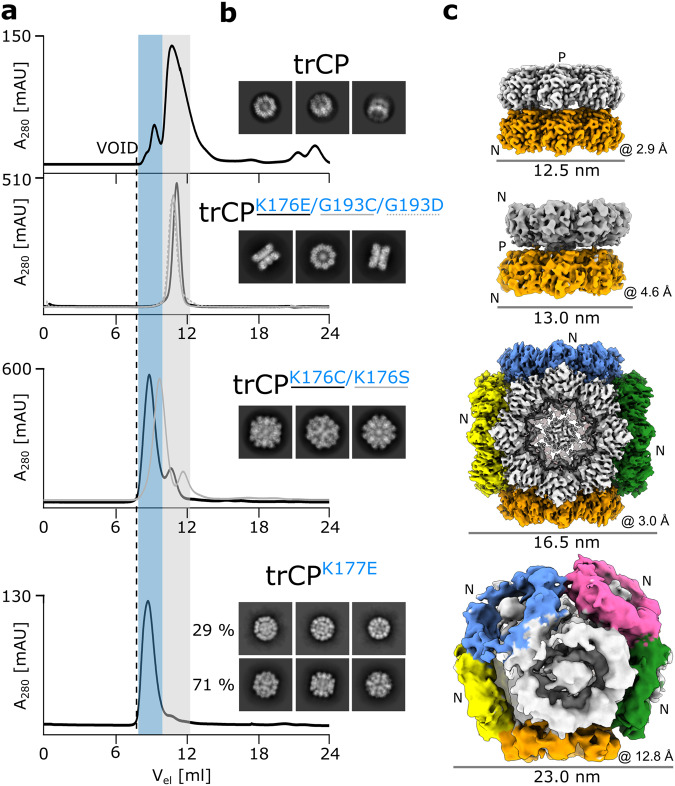
The structural diversity and tunability of the capsid proteins (CPs) of various icosahedral and rod-shaped viruses have been well studied and exploited in the development of smart hybrid nanoparticles. However, the potential of CPs of the wide-spread flexuous filamentous plant viruses remains to be explored. Here, we show that we can control the shape, size, RNA encapsidation ability, symmetry, stability and surface functionalization of nanoparticles through structure-based design of CP from potato virus Y (PVY). We provide high-resolution insight into CP-based self-assemblies, ranging from large polymorphic or monomorphic filaments to smaller annular, cubic or spherical particles. Furthermore, we show that we can prevent CP self-assembly in bacteria by fusion with a cleavable protein, enabling controlled nanoparticle formation in vitro. Understanding the remarkable structural diversity of PVY CP not only provides possibilities for the production of biodegradable nanoparticles, but may also advance future studies of CP's polymorphism in a biological context.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Complementation of the movement-deficient mutations in potato virus X: potyvirus coat protein mediates cell-to-cell trafficking of C-terminal truncation but not deletion mutant of potexvirus coat protein.Virology. 2000 Apr 25;270(1):31-42. doi: 10.1006/viro.2000.0246. Virology. 2000. PMID: 10772977
-
Potato virus Y-like particles as a new carrier for the presentation of foreign protein stretches.Mol Biotechnol. 2012 Oct;52(2):129-39. doi: 10.1007/s12033-011-9480-9. Mol Biotechnol. 2012. PMID: 22167454
-
Helical capsids of plant viruses: architecture with structural lability.J Gen Virol. 2016 Aug;97(8):1739-1754. doi: 10.1099/jgv.0.000524. Epub 2016 Jun 15. J Gen Virol. 2016. PMID: 27312096 Review.
-
Structural basis for the multitasking nature of the potato virus Y coat protein.Sci Adv. 2019 Jul 17;5(7):eaaw3808. doi: 10.1126/sciadv.aaw3808. eCollection 2019 Jul. Sci Adv. 2019. PMID: 31328164 Free PMC article.
-
Potyviral coat protein and genomic RNA: A striking partnership leading virion assembly and more.Adv Virus Res. 2020;108:165-211. doi: 10.1016/bs.aivir.2020.09.001. Epub 2020 Sep 18. Adv Virus Res. 2020. PMID: 33837716 Review.
Cited by
-
Production of Plant Virus-Derived Hybrid Nanoparticles Decorated with Different Nanobodies.ACS Nano. 2024 Dec 17;18(50):33890-33906. doi: 10.1021/acsnano.4c07066. Epub 2024 Dec 2. ACS Nano. 2024. PMID: 39622501 Free PMC article.
-
High-Fidelity In Vitro Packaging of Diverse Synthetic Cargo into Encapsulin Protein Cages.Angew Chem Int Ed Engl. 2025 Jun 2;64(23):e202422459. doi: 10.1002/anie.202422459. Epub 2025 May 7. Angew Chem Int Ed Engl. 2025. PMID: 40139971 Free PMC article.
-
A large-scale curated and filterable dataset for cryo-EM foundation model pre-training.Sci Data. 2025 Jun 7;12(1):960. doi: 10.1038/s41597-025-05179-2. Sci Data. 2025. PMID: 40483273 Free PMC article.
-
3D-Printed Self-Assembling Helical Models for Exploring Viral Capsid Structures.Biomimetics (Basel). 2024 Dec 16;9(12):763. doi: 10.3390/biomimetics9120763. Biomimetics (Basel). 2024. PMID: 39727767 Free PMC article.
-
Structural Basis for Alternative Self-Assembly Pathways Leading to Different Human Immunodeficiency Virus Capsid-Like Nanoparticles.ACS Nano. 2024 Oct 8;18(40):27465-27478. doi: 10.1021/acsnano.4c07948. Epub 2024 Sep 27. ACS Nano. 2024. PMID: 39329375
References
-
- Carstens, E. B. Introduction to virus taxonomy in virus taxonomy. In Ninth Report of the International Committee on Taxonomy of Viruses (ed. King, A. M. Q., Adams, M. J., Carstens, E. B. & Lefkowitz, E.) 3–20 (Elsevier Inc, 2012).
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
