

CRISPR Activation Reverses Haploinsufficiency and Functional Deficits Caused by TTN Truncation Variants
- PMID: 38235591
- PMCID: PMC11031707
- DOI: 10.1161/CIRCULATIONAHA.123.063972
CRISPR Activation Reverses Haploinsufficiency and Functional Deficits Caused by TTN Truncation Variants
Abstract
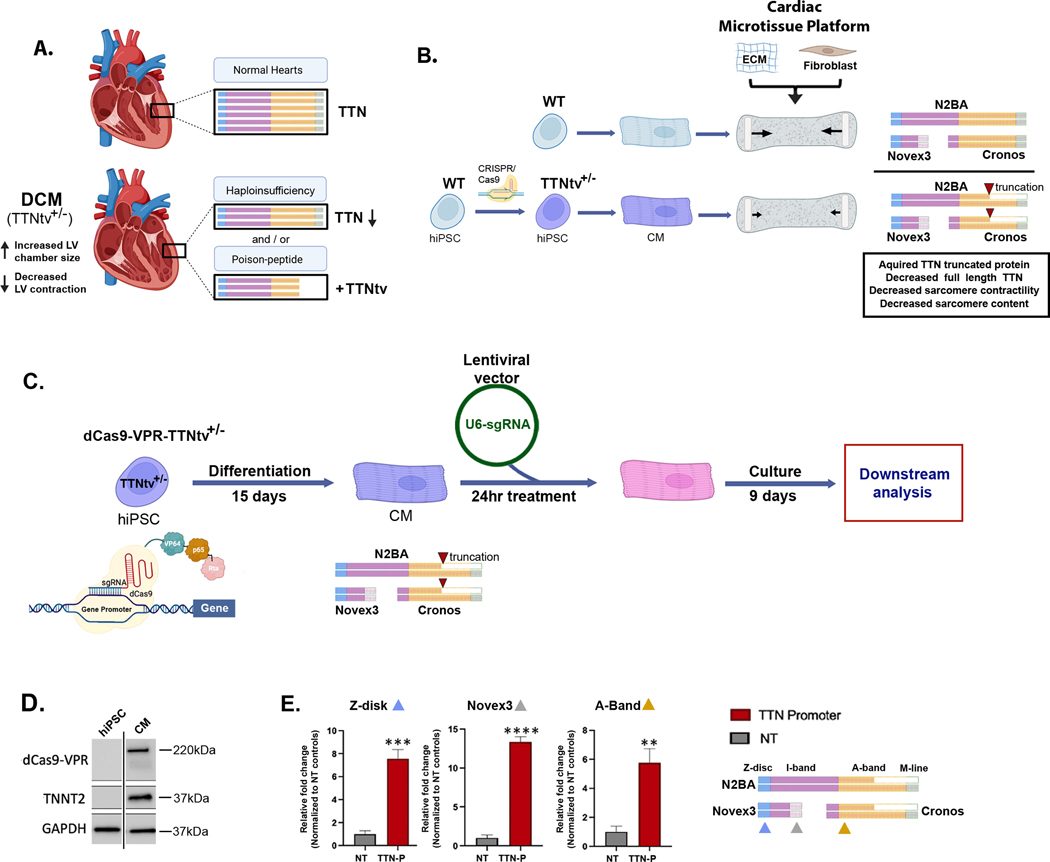
Background: TTN truncation variants (TTNtvs) are the most common genetic lesion identified in individuals with dilated cardiomyopathy, a disease with high morbidity and mortality rates. TTNtvs reduce normal TTN (titin) protein levels, produce truncated proteins, and impair sarcomere content and function. Therapeutics targeting TTNtvs have been elusive because of the immense size of TTN, the rarity of specific TTNtvs, and incomplete knowledge of TTNtv pathogenicity.
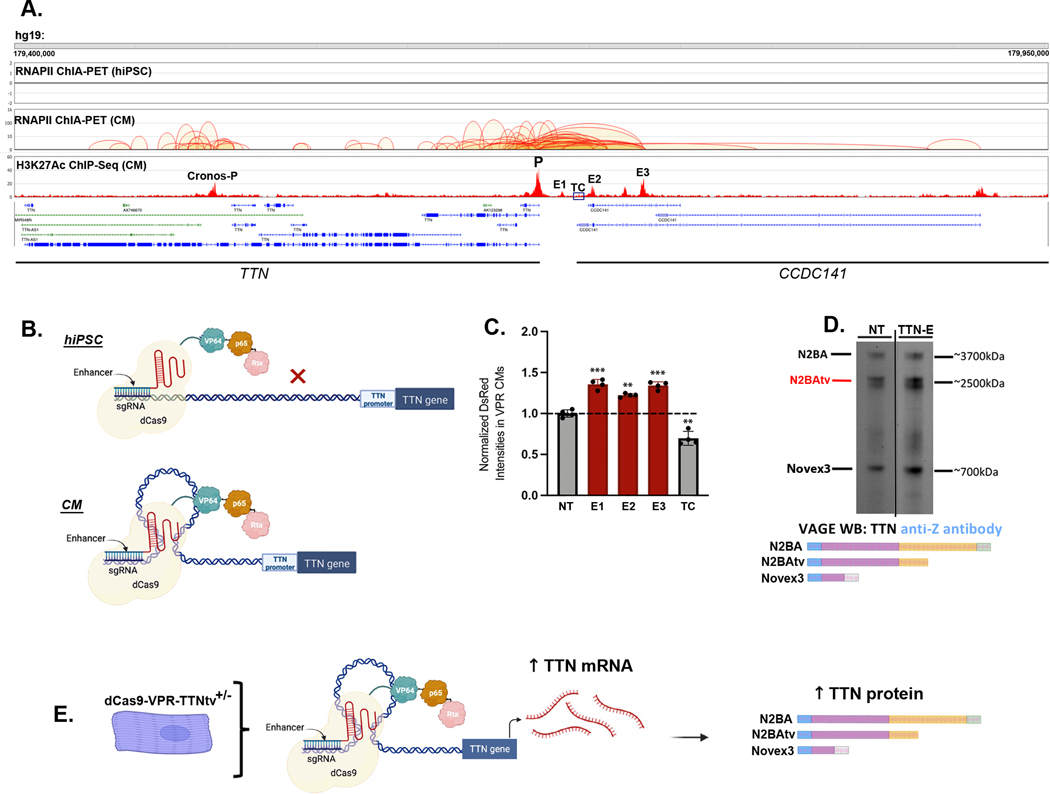
Methods: We adapted CRISPR activation using dCas9-VPR to functionally interrogate TTNtv pathogenicity and develop a therapeutic in human cardiomyocytes and 3-dimensional cardiac microtissues engineered from induced pluripotent stem cell models harboring a dilated cardiomyopathy-associated TTNtv. We performed guide RNA screening with custom TTN reporter assays, agarose gel electrophoresis to quantify TTN protein levels and isoforms, and RNA sequencing to identify molecular consequences of TTN activation. Cardiomyocyte epigenetic assays were also used to nominate DNA regulatory elements to enable cardiomyocyte-specific TTN activation.
Results: CRISPR activation of TTN using single guide RNAs targeting either the TTN promoter or regulatory elements in spatial proximity to the TTN promoter through 3-dimensional chromatin interactions rescued TTN protein deficits disturbed by TTNtvs. Increasing TTN protein levels normalized sarcomere content and contractile function despite increasing truncated TTN protein. In addition to TTN transcripts, CRISPR activation also increased levels of myofibril assembly-related and sarcomere-related transcripts.
Conclusions: TTN CRISPR activation rescued TTNtv-related functional deficits despite increasing truncated TTN levels, which provides evidence to support haploinsufficiency as a relevant genetic mechanism underlying heterozygous TTNtvs. CRISPR activation could be developed as a therapeutic to treat a large proportion of TTNtvs.
Keywords: cardiomyopathy, dilated; clustered regularly interspaced short palindromic repeats; connectin; myocytes, cardiac.
Conflict of interest statement
Figures



References
-
- Hershberger RE, Jordan E. Dilated Cardiomyopathy Overview. In: Adam MP, Everman DB, Mirzaa GM, Pagon RA, Wallace SE, Bean LJH, Gripp KW, Amemiya A, eds. GeneReviews((R)). Seattle (WA); 1993. - PubMed
-
- Roberts AM, Ware JS, Herman DS, Schafer S, Baksi J, Bick AG, Buchan RJ, Walsh R, John S, Wilkinson S, et al. Integrated allelic, transcriptional, and phenomic dissection of the cardiac effects of titin truncations in health and disease. Sci Transl Med. 2015;7:270ra276. doi: 10.1126/scitranslmed.3010134 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
