

Novel GNE missense variants impair de novo sialylation and cause defective angiogenesis in the developing brain in mice
- PMID: 38237079
- PMCID: PMC10879683
- DOI: 10.1182/bloodadvances.2023011490
Novel GNE missense variants impair de novo sialylation and cause defective angiogenesis in the developing brain in mice
Abstract
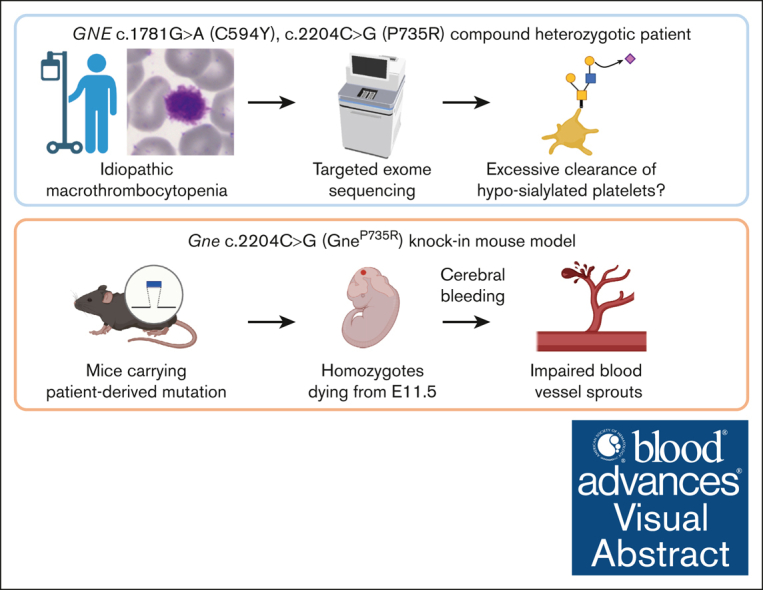
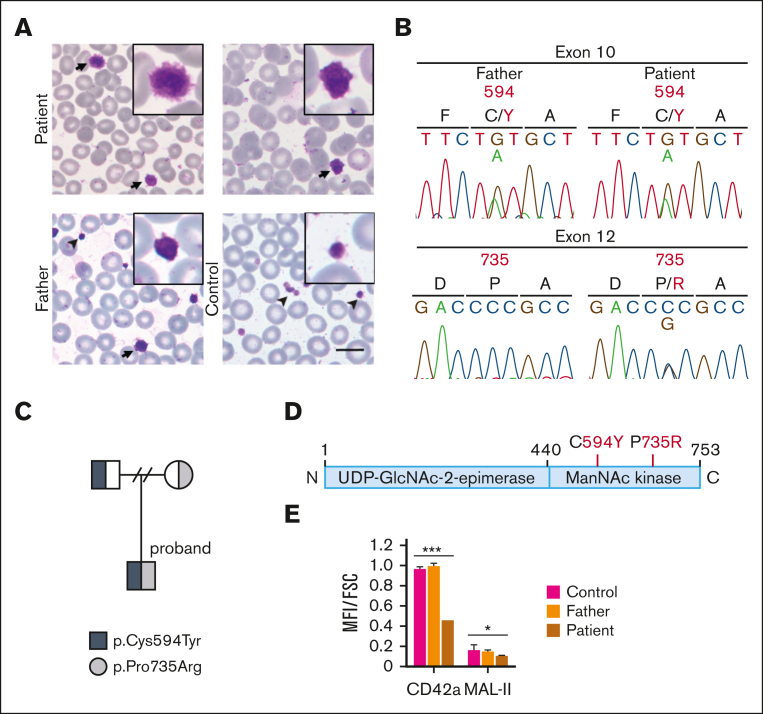
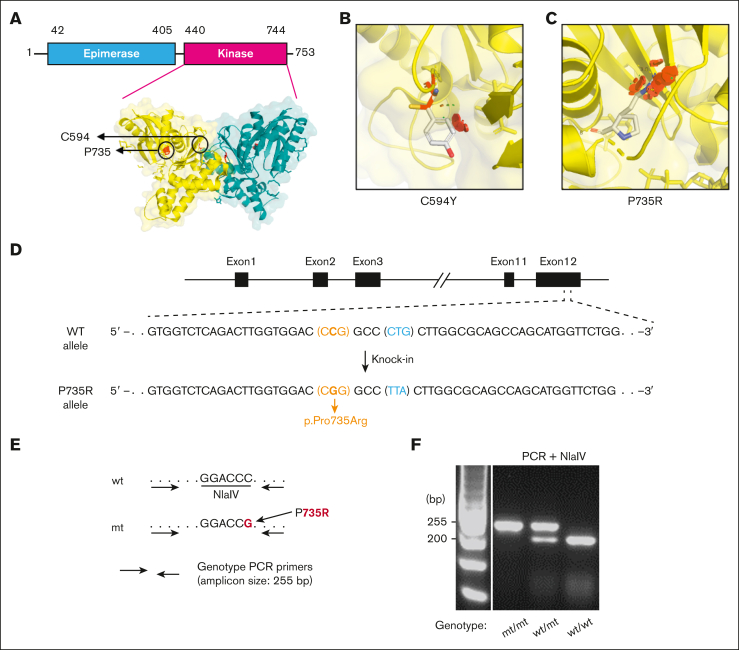
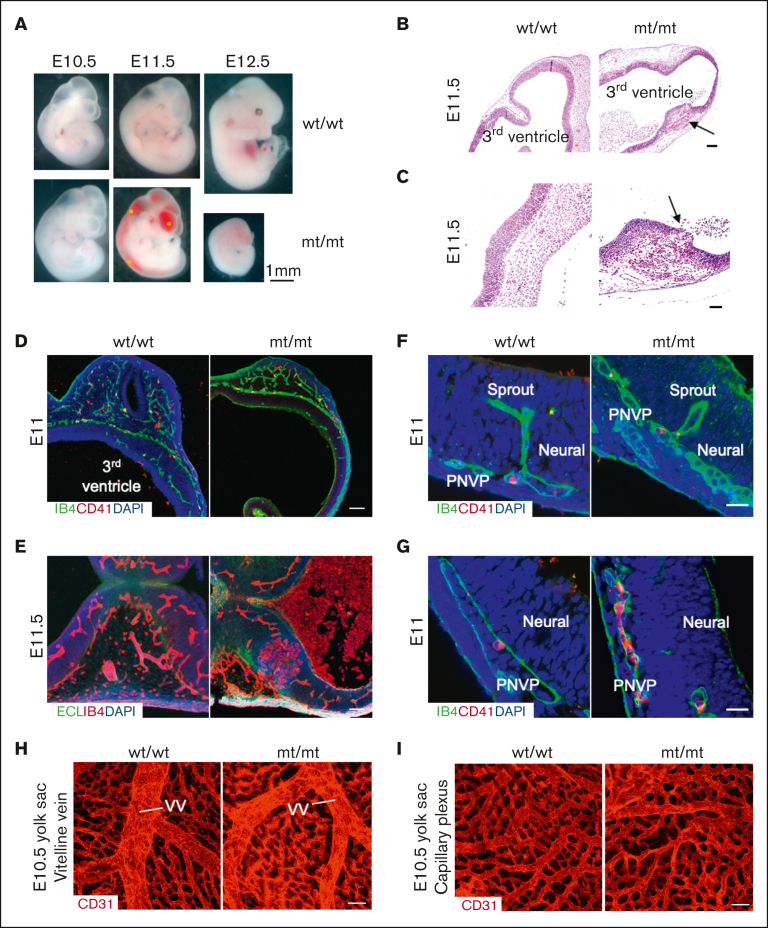
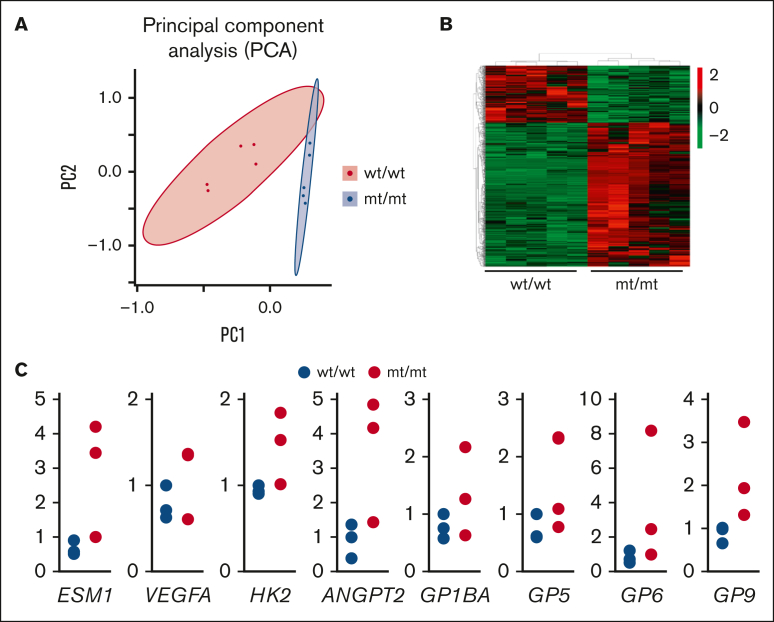
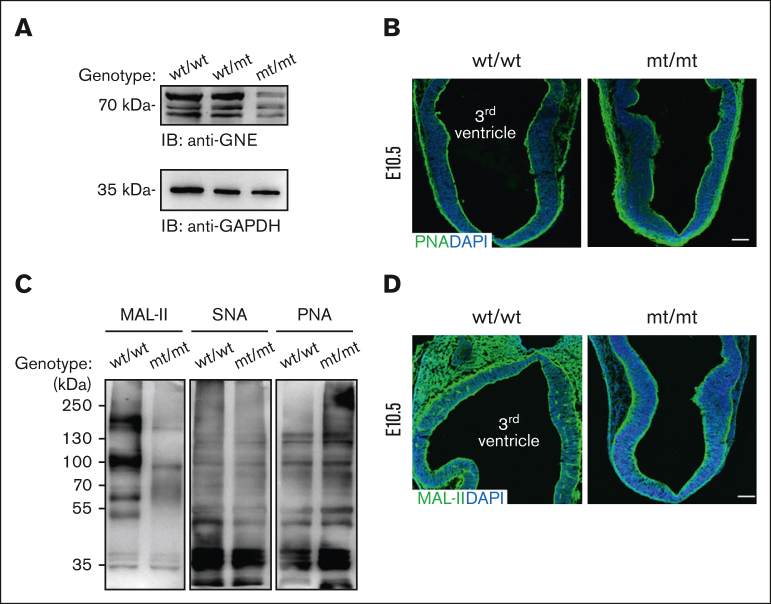
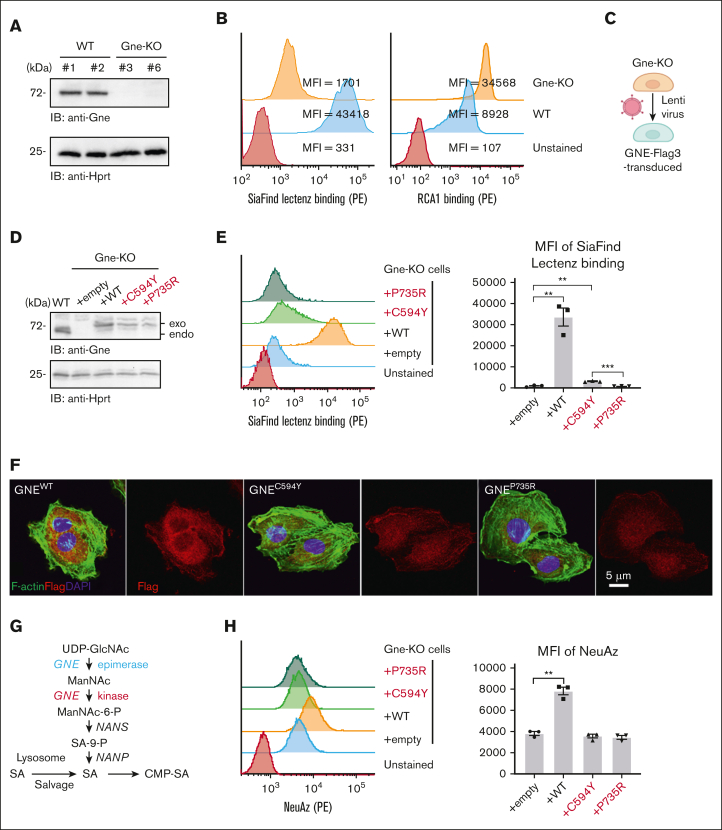
Glucosamine (UDP-N-acetyl)-2-epimerase and N-acetylmannosamine (ManNAc) kinase (GNE) is a cytosolic enzyme in de novo sialic acid biosynthesis. Congenital deficiency of GNE causes an autosomal recessive genetic disorder associated with hereditary inclusion body myopathy and macrothrombocytopenia. Here, we report a pediatric patient with severe macrothrombocytopenia carrying 2 novel GNE missense variants, c.1781G>A (p.Cys594Tyr, hereafter, C594Y) and c.2204C>G (p.Pro735Arg, hereafter, P735R). To investigate the biological significance of these variants in vivo, we generated a mouse model carrying the P735R mutation. Mice with homozygous P735R mutations exhibited cerebral hemorrhages as early as embryonic day 11 (E11), which subsequently progressed to large hemorrhages in the brain and spinal cord, and died between E11.5 and E12.5. Defective angiogenesis such as distended vascular sprouts were found in neural tissues and embryonic megakaryocytes were abnormally accumulated in the perineural vascular plexus in mutant mouse embryos. Furthermore, our in vitro experiments indicated that both C594Y and P735R are loss-of-function mutations with respect to de novo sialic acid biosynthesis. Overall, this study reveals a novel role for GNE-mediated de novo sialic acid biosynthesis in mouse embryonic angiogenesis.
© 2024 by The American Society of Hematology. Licensed under Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0), permitting only noncommercial, nonderivative use with attribution. All other rights reserved.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
Similar articles
-
Safety, pharmacokinetics and sialic acid production after oral administration of N-acetylmannosamine (ManNAc) to subjects with GNE myopathy.Mol Genet Metab. 2017 Sep;122(1-2):126-134. doi: 10.1016/j.ymgme.2017.04.010. Epub 2017 Apr 26. Mol Genet Metab. 2017. PMID: 28641925 Free PMC article. Clinical Trial.
-
Lec3 Chinese hamster ovary mutants lack UDP-N-acetylglucosamine 2-epimerase activity because of mutations in the epimerase domain of the Gne gene.J Biol Chem. 2003 Dec 26;278(52):53045-54. doi: 10.1074/jbc.M309967200. Epub 2003 Oct 15. J Biol Chem. 2003. PMID: 14561743
-
Uridine diphosphate-N-acetylglucosamine-2-epimerase/N-acetylmannosamine kinase deletion in mice leads to lethal intracerebral hemorrhage during embryonic development.Glycobiology. 2021 Dec 18;31(11):1478-1489. doi: 10.1093/glycob/cwab069. Glycobiology. 2021. PMID: 34224569
-
Regulation and pathophysiological implications of UDP-GlcNAc 2-epimerase/ManNAc kinase (GNE) as the key enzyme of sialic acid biosynthesis.Biol Chem. 2009 Jul;390(7):591-9. doi: 10.1515/BC.2009.073. Biol Chem. 2009. PMID: 19426133 Review.
-
[Development of therapy for distal myopathy with rimmed vacuoles].Rinsho Shinkeigaku. 2009 Nov;49(11):852-5. doi: 10.5692/clinicalneurol.49.852. Rinsho Shinkeigaku. 2009. PMID: 20030229 Review. Japanese.
Cited by
-
Induced Muscle and Liver Absence of Gne in Postnatal Mice Does Not Result in Structural or Functional Muscle Impairment.J Neuromuscul Dis. 2024;11(5):905-917. doi: 10.3233/JND-240056. J Neuromuscul Dis. 2024. PMID: 38875046 Free PMC article.
References
-
- Hinderlich S, Stäsche R, Zeitler R, Reutter W. A bifunctional enzyme catalyzes the first two steps in N- acetylneuraminic acid biosynthesis of rat liver. Purification and characterization of UDP-N-acetylglucosamine 2-epimerase/N-acetylmannosamine kinase. J Biol Chem. 1997;272(39):24313–24318. - PubMed
-
- Tanner ME. The enzymes of sialic acid biosynthesis. Bioorg Chem. 2005;33(3):216–228. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
