

The transcriptional cofactor Tle3 reciprocally controls effector and central memory CD8+ T cell fates
- PMID: 38238608
- PMCID: PMC10916363
- DOI: 10.1038/s41590-023-01720-w
The transcriptional cofactor Tle3 reciprocally controls effector and central memory CD8+ T cell fates
Erratum in
-
Publisher Correction: The transcriptional cofactor Tle3 reciprocally controls effector and central memory CD8+T cell fates.Nat Immunol. 2024 Apr;25(4):717. doi: 10.1038/s41590-024-01780-6. Nat Immunol. 2024. PMID: 38347084 No abstract available.
Abstract
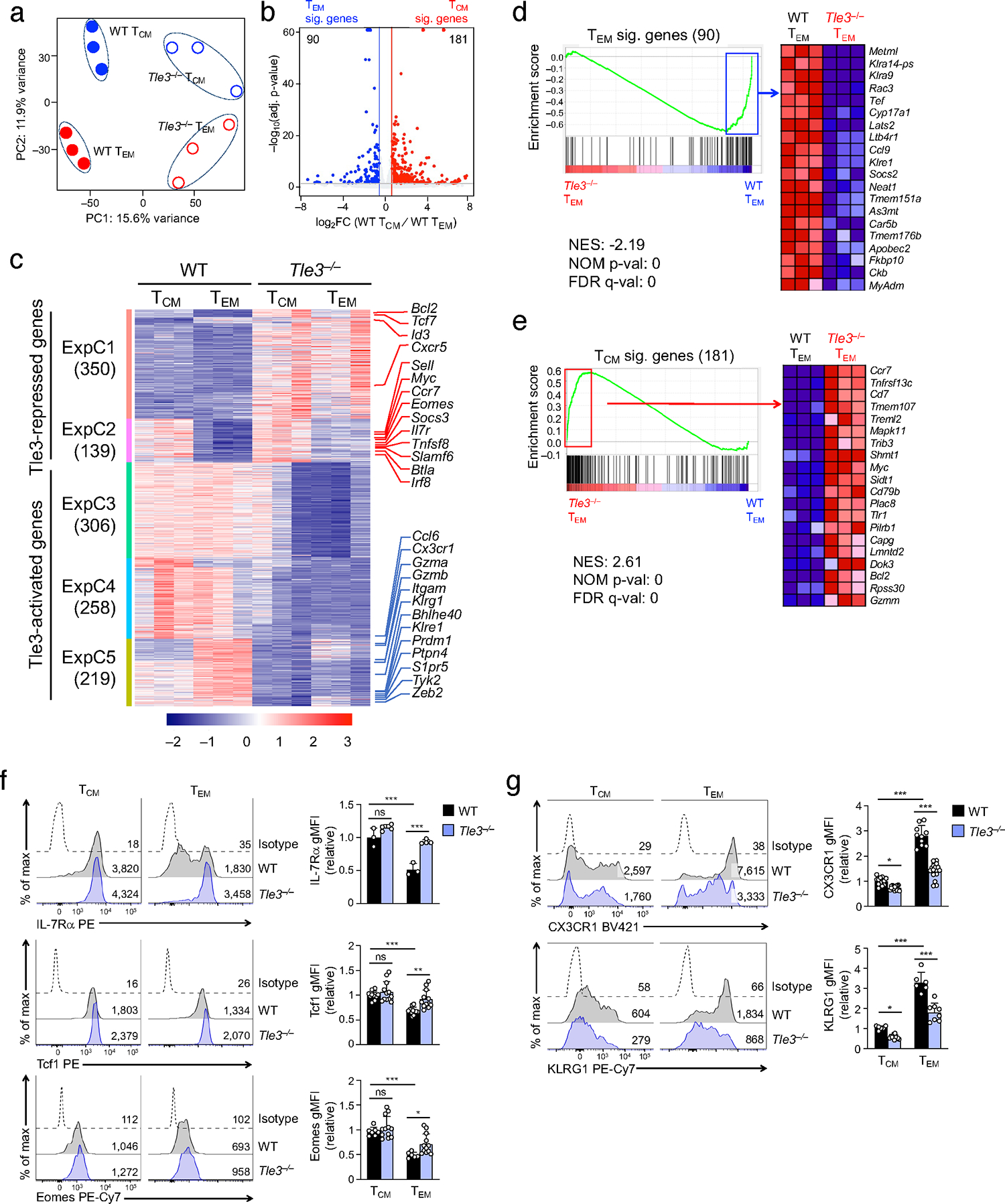
Antigen-experienced CD8+ T cells form effector and central memory T cells (TEM and TCM cells, respectively); however, the mechanism(s) controlling their lineage plasticity remains incompletely understood. Here we show that the transcription cofactor Tle3 critically regulates TEM and TCM cell fates and lineage stability through dynamic redistribution in antigen-responding CD8+ T cell genome. Genetic ablation of Tle3 promoted CD8+ TCM cell formation at the expense of CD8+ TEM cells. Lineage tracing showed that Tle3-deficient CD8+ TEM cells underwent accelerated conversion into CD8+ TCM cells while retaining robust recall capacity. Tle3 acted as a coactivator for Tbet to increase chromatin opening at CD8+ TEM cell-characteristic sites and to activate CD8+ TEM cell signature gene transcription, while engaging Runx3 and Tcf1 to limit CD8+ TCM cell-characteristic molecular features. Thus, Tle3 integrated functions of multiple transcription factors to guard lineage fidelity of CD8+ TEM cells, and manipulation of Tle3 activity could favor CD8+ TCM cell production.
© 2024. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Figures














References
-
- Sallusto F, Lenig D, Forster R, Lipp M & Lanzavecchia A Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 401, 708–712 (1999). - PubMed
-
- Wherry EJ et al. Lineage relationship and protective immunity of memory CD8 T cell subsets. Nature immunology 4, 225–234 (2003). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
