

Lipid vesicle-based molecular robots
- PMID: 38239102
- PMCID: PMC10898420
- DOI: 10.1039/d3lc00860f
Lipid vesicle-based molecular robots
Abstract
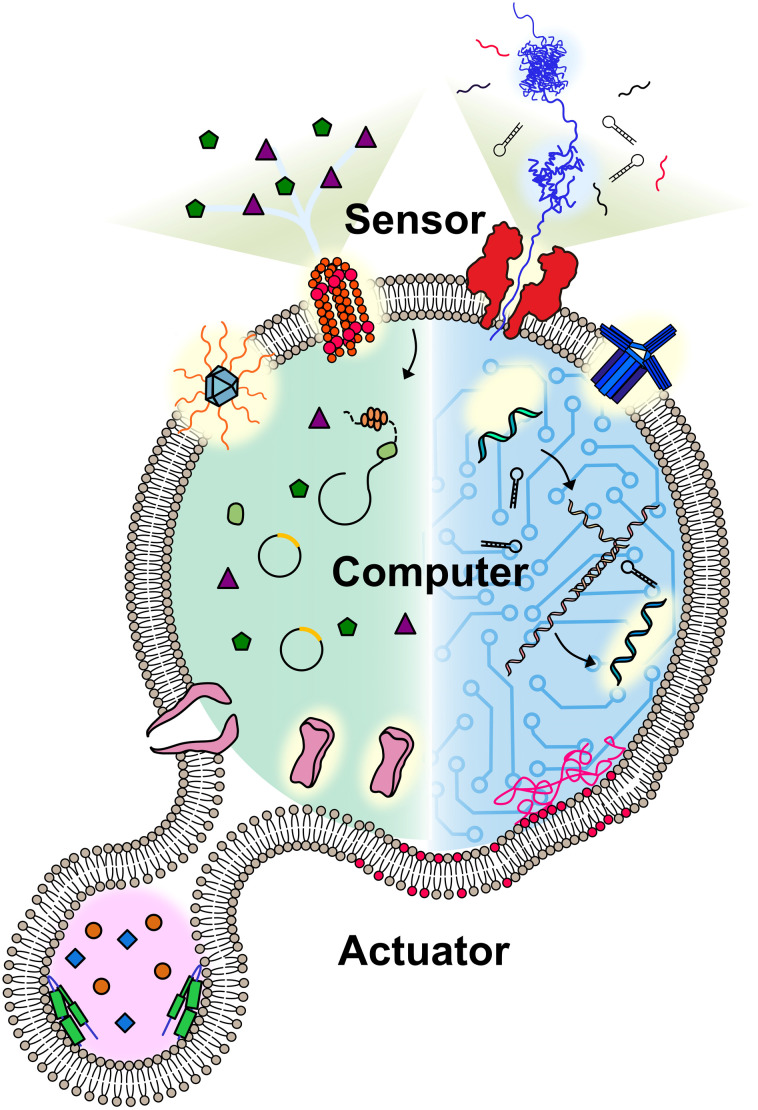
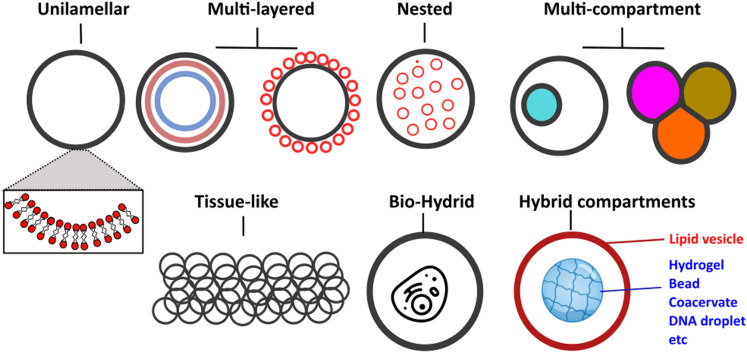
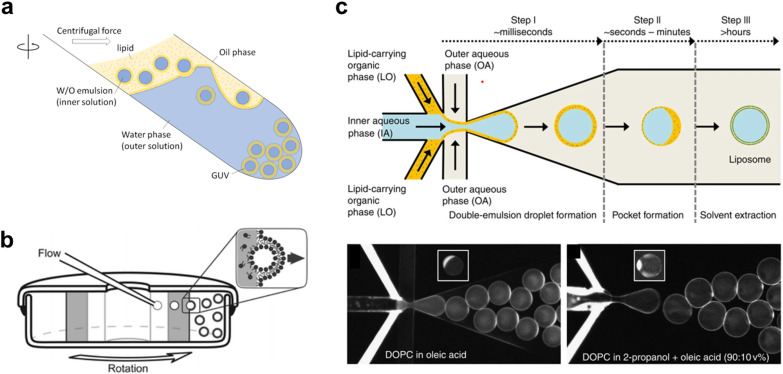
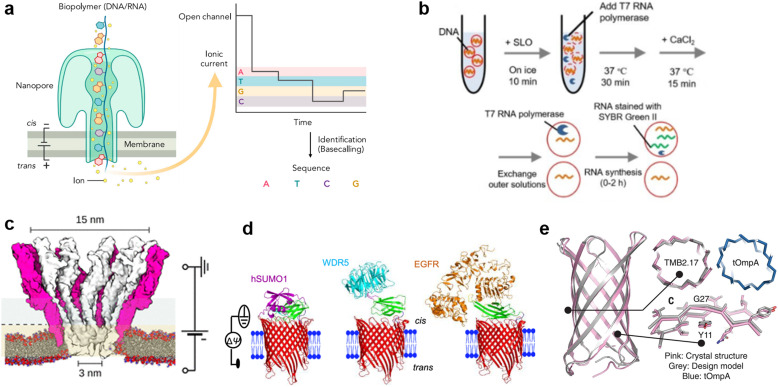
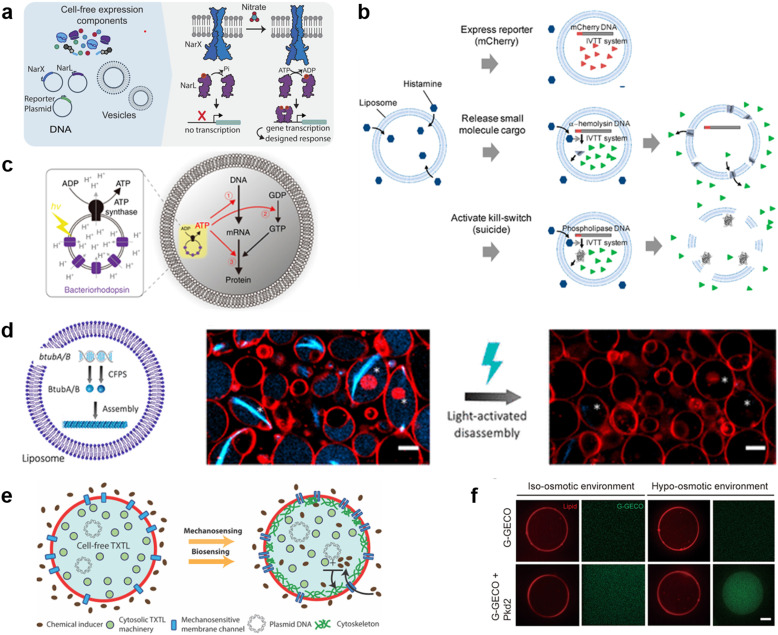
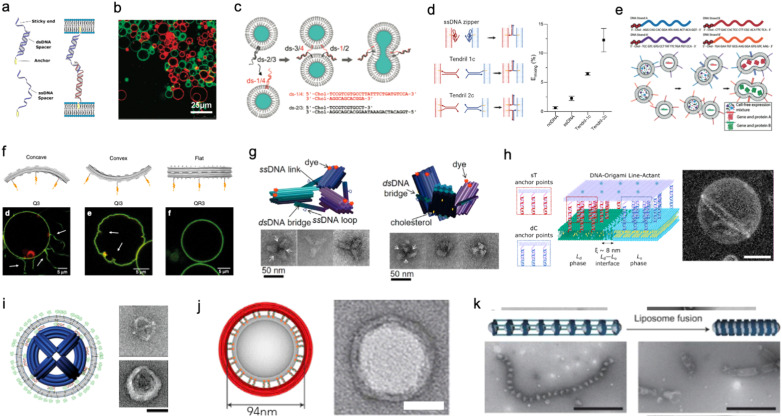
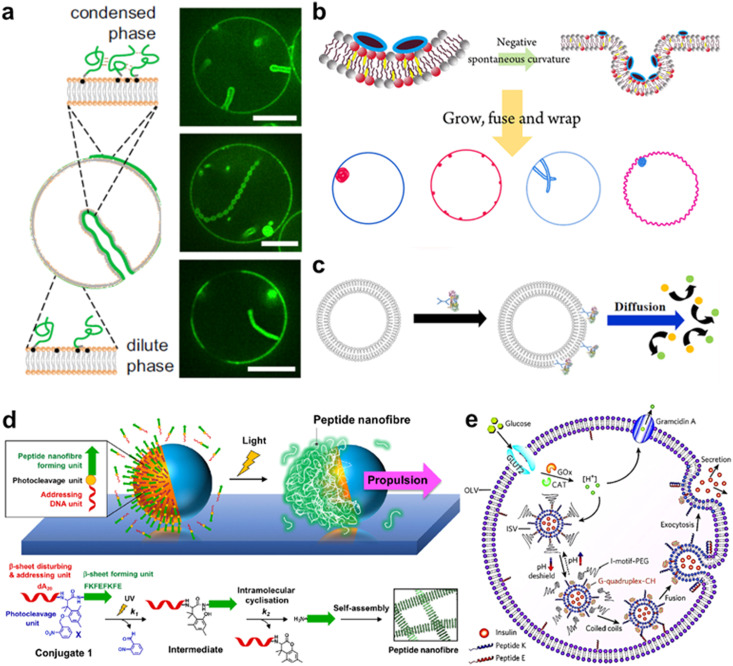
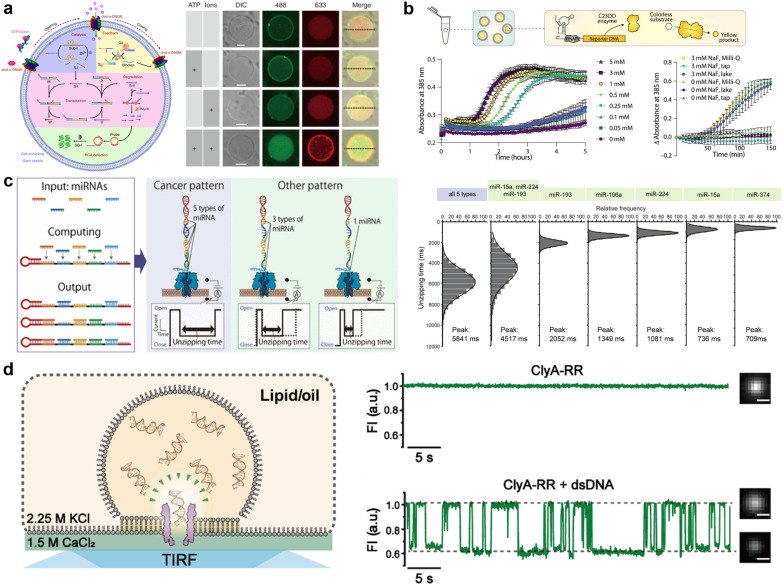
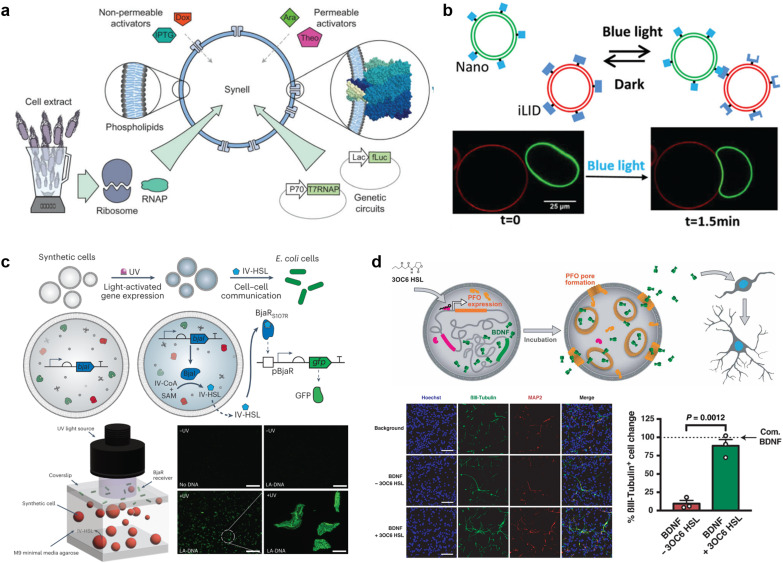
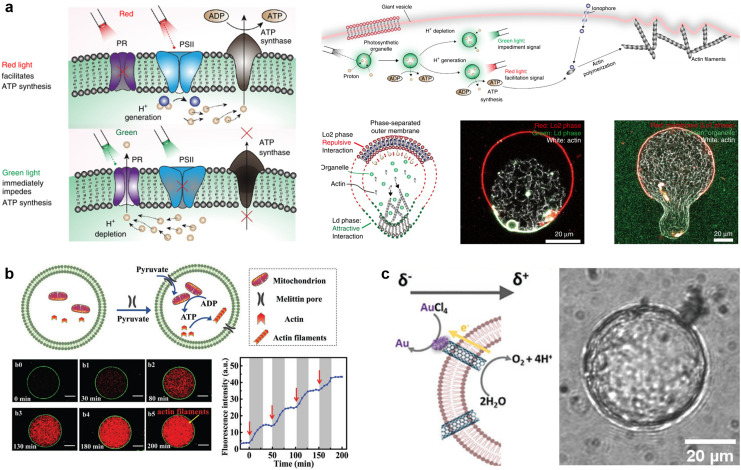
A molecular robot, which is a system comprised of one or more molecular machines and computers, can execute sophisticated tasks in many fields that span from nanomedicine to green nanotechnology. The core parts of molecular robots are fairly consistent from system to system and always include (i) a body to encapsulate molecular machines, (ii) sensors to capture signals, (iii) computers to make decisions, and (iv) actuators to perform tasks. This review aims to provide an overview of approaches and considerations to develop molecular robots. We first introduce the basic technologies required for constructing the core parts of molecular robots, describe the recent progress towards achieving higher functionality, and subsequently discuss the current challenges and outlook. We also highlight the applications of molecular robots in sensing biomarkers, signal communications with living cells, and conversion of energy. Although molecular robots are still in their infancy, they will unquestionably initiate massive change in biomedical and environmental technology in the not too distant future.
Conflict of interest statement
There are no conflicts to declare.
Figures
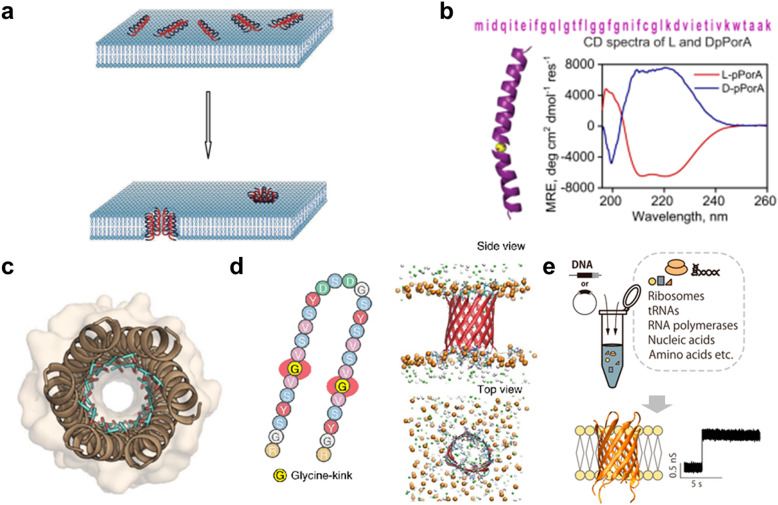
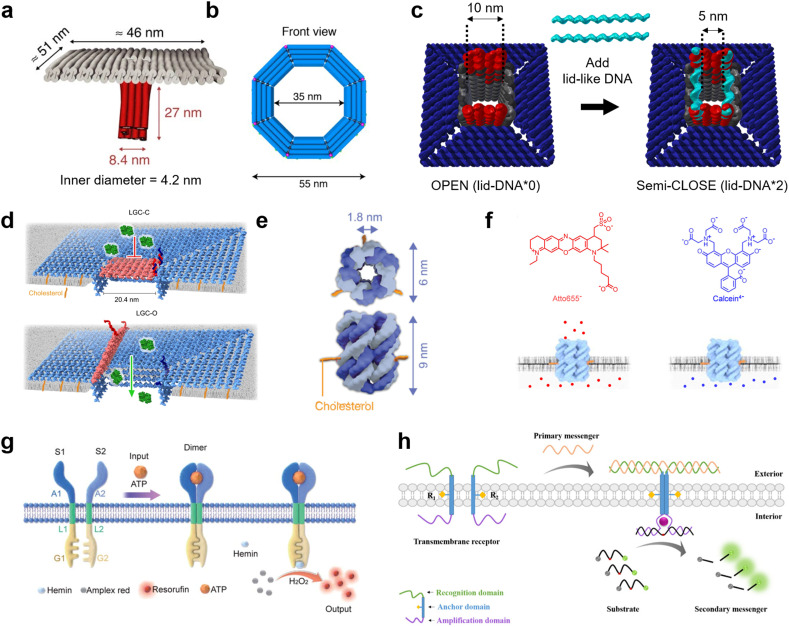
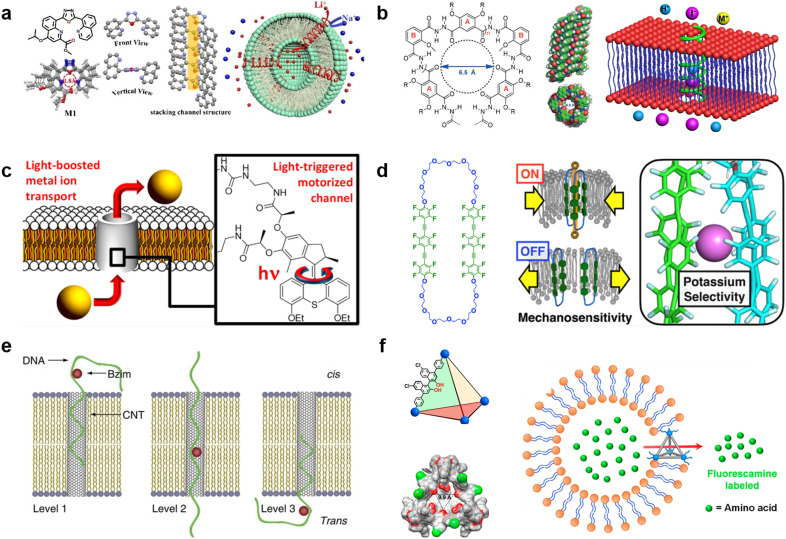
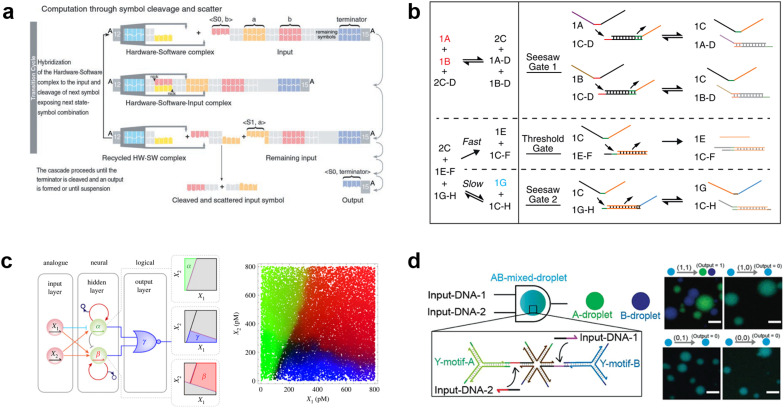
References
-
- Dietrich-Buchecker C. O. Sauvage J. P. Kintzinger J. P. Une nouvelle famille de molecules : les metallo-catenanes. Tetrahedron Lett. 1983;24:5095–5098. doi: 10.1016/S0040-4039(00)94050-4. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources